ENRIQUE RUBIO GARCIA
Jefe del Servicio de Neurocirugía Valle de Hebron
Profesor Titular de Neurocirugía
Academico de España, Portugal, European Society of Neurosurgery, Word Federation of Neurosurgery.
Investigador del I Carlos III
Veintidós tesis doctorales dirigidas
250 trabajos publicados
Presidente de la academia de Neurocirugía de Barcelona
Academico de Cadiz y Jerez de la Frontera
Acadenico de Honor de Andalucia y Cataluña
log enriquerubio.net
Carl Sagan fue un divulgador genial entre otras capacidades pero no se le puede apartar de su esposa. Lynn Margulis, la pasión por la vida
Su estudio de la evolución celular modificó nuestra percepción de la biosfera
Carl Sagan durante su intervención en el Congreso de los Estados Unidos en 1985
Lynn Margulis, la pasión por la vida
En 1985 el Congreso de los Estados Unidos, reunido en Washington, promovió un estudio sobre los gases de efecto invernadero y su efecto sobre el clima.
Y el astronomo y científico Carl Sagan, el científico,fue encargado de una conferencia que ha sido después referencial
Las grandes compañías petroleras eran completamente conocedoras del problema que para el planeta suponía seguir quemando combustibles fósiles de forma incontrolada, Y por supuesto hacían todo lo posible por evitar que este discurso llegara al gran publico y Car Sagan fue tan acertado en su charla que lleno al mundo y lo sigue llenado de los peligros de los gases invernaderos.
Un año después de esta charla, 1986 la industria petrolera accedió gracias a la campaña de Clair Patterson , a eliminar el dañino plomo de su gasolina, después de una batalla que duró varias décadas
Sagan dijo acertadamente :«Tendemos a pensar que el asunto de los gases de efecto invernadero es un problema para la gente del siglo XXI, de modo que ya se preocuparán ellos llegado el momento. Lo cierto es que si no actuamos ahora luego será demasiado tarde. Les estamos traspasando a nuestros hijos problemas extremadamente graves, cuando el momento para evitarlos – si es que en realidad pueden evitarse – es ahora».
El astrónomo y divulgador no desaprovechó la ocasión y explicó que en principio un poco de efecto invernadero es beneficioso. Tras hacer los cálculos entre la cantidad de energía solar que llega a la Tierra en cada momento, y el volumen de radiación UV que nuestro planeta rebota al espacio, la temperatura en la Tierra debería ser 30ºC más baja, y que si esto no sucedía era precisamente por las propiedades “absorbentes” de calor que muestran el CO2 y otros gases invernadero. Si nuestro planeta no atravesaba una edad del hielo perpetua era por los gases invernadero. No obstante lo dejó claro: “un poco es bueno, pero hay un equilibrio delicado”.
Para ejemplificar este delicado balance, Sagan explicó que la diferencia climática en el pasado entre las edades glaciares y las interglaciares se debieron a pequeñas variaciones en la cantidad de luz solar que alcanzaba la superficie del planeta, las cuales se producían a su vez por pequeños cambios en la órbita terrestre. Por ello, para saber más sobre el efecto de los gases invernadero en el clima es necesario estudiar las características de la Tierra en el pasado, recomienda el catedrático.
El mundo científico siguió su consejo ya que a día de hoy se han producido múltiples trabajos basados en las burbujas de aire atrapadas en testigos de hielo extraídos de glaciares y de los casquetes polares, lo que ha permitido medir las cantidades de CO2 atmosférico en el pasado.
Sagan denuncio como los gases de efecto invernadero han afectado a otros planetas, y el ejemplo de Venus es inmediato. “Un mundo similar a la Tierra en densidad, masa, situación, etc. pero con 90 veces más CO2 atmosférico que nuestro mundo, lo cual provoca temperaturas superficiales superiores a los 400ºC”.
La advertencia a los políticos de Washington sobre el papel desestabilizador que nuestra especie juega a nivel planetario no deja dudas:
“El poder de los seres humanos para afectar, controlar y cambiar el medioambiente cambia a medida que nuestra tecnología mejora. En estos momentos, claramente hemos alcanzado la etapa en la que somos capaces (tanto intencionada como inadvertidamente) de alterar de forma significativa el clima y el ecosistema a nivel global. Probablemente llevamos haciéndolo – aunque a menor escala – desde hace mucho tiempo”.
El ejemplo de Egipto en tiempos del imperio romano salió a colación durante el discurso ante el congreso. “Egipto era el granero de Roma hace dos milenios. Hoy en día es una tierra árida a poco que te alejes del Nilo, algo que seguramente provocó el ser humano por un exceso de pastoreo.
Apuntaba Sagan como dejar de quemar combustibles fósiles tiene unas implicaciones económicas tan severas que nadie se lo plantea en serio, pero hay otras muchas cosas que se pueden hacer”, invertir en otras fuentes energéticas como la solar, la fisión nuclear “segura” (lo cual es factible, añade) y a largo plazo la fusión nuclear. ¡Si amigos ecologistas, Sagan apoyaba la energía nuclear!
De igual forma Sagan no menciono los efectos toxicos de la polucion en general, que por lo menos igualan a los gases invernaderos.
El objetivo requerirá un consenso global que ahora mismo dista mucho de estar cerca. Lo esencial es adquirir una conciencia global, una visión que trascienda nuestras identificaciones exclusivas. Todos los grupos políticos y naciones deben cooperar, porque en este ‘ìnvernadero’ estamos todos juntos«.
Esta claro que la brillantes de Sagan se hizo escuchar y es efectiva, pero no solo me encanta Sagan, lo hace su esposa, que sin filantropía, habla del mundo de lo orgánico
Lynn Margulis, la pasión por la vida
Su estudio de la evolución celular modificó nuestra percepción de la biosfera
Esta Sra, que se canso de Sagan como de otros esposos, era de una vitalidad inigualable que llevó a los estudiantes de Boston a apodarla Nuestra Señora del Movimiento Perpetuo, la mezcla explosiva de su inteligencia, irreverencia científica y enorme calidez sedujo a un sinnúmero de alumnos, colegas y amigos que seguimos lamentando su muerte prematura ocurrida el pasado 22 de noviembre a los 73 años, a consecuencia de una embolia.
Enmendó la plana a Linneo y Haeckel, reclasificando todas las criaturas vivas
Es imposible separar la vida personal de Lynn Margulis de su biografía científica. A los 14 años ingresó a la Universidad de Chicago, y al concluir sus estudios de Biología tres años mas tarde se marchó a México para trabajar en un proyecto de etnobotánica al lado de Oscar Lewis, el legendario antropólogo de la pobreza que estudiaba la vida cotidiana de Tepoztlán.
Tenia Lynn amor por todas las variantes del castellano que había comenzado a conocer cuando se aventuraba con audacia adolescente a los barrios bajos de su natal Chicago.
Cuando regresó 30 años después las ancianas salían de sus casas a abrazarla, presentarle a sus hijos y nietos e invitarnos a comer sentados en el suelo en torno al fogón.
En EEUU se casó con Carl Sagan, tuvo sus primeros dos hijos y terminó su doctorado. Se divorció de Sagan y se casó con Nick Margulis, un químico estadounidense, y tuvo dos hijos mas, pero se volvió a separar porque no quiso cuidarle el fogón a Carl Sagan ni a ningún otro.
Haciendo gala de una intuición excepcional y de un conocimiento formidable de la biología, Lynn Margulis no tardó en darse cuenta del papel central que la simbiosis jugó en el origen de las células nucleadas de plantas, hongos, animales y un sinnúmero de microorganismos. Así, mientras algunos miraban hacia el DNA, ella comenzó a asomarse hacia el planeta, lo que le permitió describir la relación entre algunos procesos centrales de la evolución biológica con la transformación de la atmósfera, los sedimentos y la hidrosfera de la Tierra misma. Algunas veces se equivocó, pero sus aciertos modificaron para siempre nuestra percepción de la biosfera.
Su hijo Dorion Sagan afirmó que trataba a sus alumnos como si fueran sus hijos, y a sus hijos como si fueran sus alumnos.
Cuando abarca el origen simbiótico de la células nucleadas, y reclasifica todas las criaturas visibles e invisibles, a las que agrupó en cinco grandes reinos.
Los microbios eran sus preferidos, y en su cartera llevaba las fotografías de sus protistas preferidos al lado de las imágenes de sus hijos y nietos.
Escritora prolífica y de un enorme refinamiento intelectual, hizo de la promoción de la visión secular de la evolución uno de sus empeños fundamentales. Fue una amiga leal, una maestra insuperable y una colega generosa, que comencé a comprender y querer mas el día en que durante una conferencia descubrí en su mirada el candor de una niña frágil pero armada de una curiosidad y una valentía irrefrenables, lista para adentrarse a lo desconocido, como lo ha hecho ahora y para siempre.
La figura de estos dos intelectuales me apasiona y me hace pensar que pese a su unión , les interesaban cosas diferentes, a ella, la vida y a Carl la publicidad, pero ambos lo hacían bien y consiguieron. entusiasmarnos
Bibliografia
Antonio Lazcano Araujo catedrático de Origen de la Vida en la Universidad Nacional Autónoma de México (UNAM).
La Organización Mundial de la Salud (OMS) decidió en mayo de 2021 recurrir al alfabeto griego para nombrar a las nuevas variantes del coronavirus. Lo hizo para evitar la estigmatización asociada al modelo que se había ido siguiendo hasta entonces: nombrar a las variantes por el país en el que emergían.
El 26 de noviembre de 2021, la OMS, siguiendo el consejo del Grupo Consultivo Técnico sobre la Evolución del Virus SARS-CoV-2- en inglés, clasificó la variante B.1.1.529 de este virus como variante preocupante. Además, decidió denominarla con la letra griega ómicron. La decisión de considerar preocupante esta variante se basó en la evidencia presentada al Grupo Consultivo Técnico, que indica que presenta varias mutaciones que podrían afectar a las características del virus, por ejemplo, la facilidad para propagarse o la gravedad de los síntomas que causa.
En Sudáfrica y en todo el mundo, los investigadores están realizando estudios para conocer mejor muchos aspectos de la variante ómicron y continuarán informando de los resultados que obtengan a medida que estén disponibles.
No está claro si, con respecto a otras variantes, como la delta, la variante ómicron es más transmisible. El número de personas que han dado positivo en las pruebas de detección ha aumentado en las zonas de Sudáfrica
Se desconoce si el cuadro clínico de la infección por la variante ómicron es más grave que el ocasionado por otras variantes, incluida la delta. De acuerdo con los datos preliminares, las tasas de hospitalización en Sudáfrica van en aumento, pero ello podría deberse a que hay más personas que han contraído la infección y no necesariamente a que se hayan infectado por la variante ómicron.
No hay datos que indiquen que los síntomas asociados con esta variante sean distintos a los ocasionados por otras variantes.
Los datos preliminares indican que, en comparación con otras variantes preocupantes, el riesgo de reinfección por la variante ómicron puede ser mayor (es decir, las personas que han tenido COVID-19 antes podrían infectarse de nuevo por esta variante con más facilidad). De todos modos, la información es todavía limitada.
La OMS trabaja con sus asociados técnicos para conocer los efectos que pueda tener esta variante en las medidas adoptadas para combatir la enfermedad, incluidas las vacunas. Todas las vacunas utilizadas actualmente continúan previniendo con mucha eficacia los síntomas graves y la muerte por COVID-19.
Las pruebas de PCR que se vienen usando ampliamente siguen detectando la infección por las distintas variantes del virus, incluida la ómicron. La eficacia de detección rápida de antígenos estas en estudio.
La OMS está coordinando a un gran número de investigadores de todo el mundo para conocer mejor la variante ómicron.
Los países deberían continuar aplicando las medidas de salud pública de eficacia demostrada para reducir de forma general la circulación del virus causante de la COVID-19, basándose en el análisis de los riesgos y en un enfoque científico.
Es fundamental reducir las desigualdades que afectan al acceso a las vacunas contra la COVID-19 para asegurarnos de que, en todos los países, se administran la primera y la segunda dosis de las vacunas a todos los grupos vulnerables de la población, incluidos los trabajadores de la salud y las personas mayores, además de ofrecerles de forma equitativa las pruebas diagnósticas y los tratamientos que necesitan.
Las enfermedades en general en los tiempos que vivimos, aunque tienen un alto contenido en organicidad, sin duda alguna el enfermo aporta un componente psiquico, que lo asusta y de manera reiterada y diaria.
Y también los medios de comunicación sin duda alguna aportan una nota de gravedad. Nos asustan cada dia.
Es necesario tener cuidado con las noticias sobre el nuevo tipo de coronavirus el ómicron ya que mientras las noticias de la televisión quitan hierro al nuevo virus según acabó de escuchar, otras noticias en Internet, lo ponen peligroso y menos sensibles a las vacuna.
Acabo de escuchar en las noticias de la televisión que hay muchos casos repartidos por el mundo de la nueva variedad del coronavirus, pero no hay ningúno grave.
De igual forma en estas noticias que proporciona Internet dicen que el nuevo virus no es demasiado sensible a la vacuna que estamos utilizando.
Moderna cree que las actuales vacunas serán “mucho menos eficaces” contra la nueva variante
Los ejecutivos de los principales fabricantes de vacunas anticiparon este martes opiniones sobre el efecto de ómicron, la más reciente mutación del coronavirus, sobre las personas vacunadas.
El consejero delegado de Moderna, Stéphane Bancel, ha afirmado que cree que las actuales vacunas contra el Covid-19 serán “mucho menos eficaces” contra la nueva variante. En contraste, Ugur Sahin, consejero delegado de BioNTech, indicó que cree que su vacuna ofrecerá una “fuerte protección” contra la nueva variante. Ambas empresas se encuentran haciendo estudios para evaluar el efecto de ómicron, por lo que los ejecutivos han aclarado que pronto tendrán respuestas concretas, en las próximas semanas.
En la sesión de este martes de la Bolsa de Nueva York, las acciones de Moderna cayeron alrededor de un 6,5%. Los títulos de BioNTech también cayeron inicialmente en la Bolsa de Fráncfort y en Wall Street, afectadas por el comentario pesimista de Moderna, pero recuperaron valor tras las declaraciones de Sahin. Por otro lado, los títulos de Pfizer, socio de BioNTech, subieron alrededor de un 2%.
La Universidad de Oxford se ha alineado con BioNTech y ha afirmado que “no hay evidencia de que la nueva variante afecte la efectividad de las vacunas”. La entidad inglesa, que junto al laboratorio AstraZeneca desarrolló otra de las vacunas, ha asegurado también que ya se encuentra haciendo estudios en países como Botsuana para probar la efectividad de sus dosis.
La Agencia Europea del Medicamento (EMA), ha señalado, por su parte, que cuenta con planes para acelerar la adaptación de las vacunas a la nueva variante. La directora de la Agencia Europea del Medicamento, Emer Cooke, ha indicado que, si fuera necesario, el proceso podría tardar entre tres y cuatro meses.
Asimismo, la farmacéutica Regeneron Pharmaceutical ha afirmado que su fármaco de anticuerpos Covid-19 también podría ser menos efectivo contra ómicron. En este sentido, la empresa ha explicado que las mutaciones podrían afectar la inmunidad adquirida por anticuerpos monoclonales.
Es necesario tener paciencia a la hora de publicar resultados que no están verificados, porque la gente sufre con las malas noticias.
“La gente se asusta con las malas noticias”. Por favor divulgadores, sean caritativos
Informadores
ALBERTO FERNANDEZ HORCAJO
Otro bluff como la varíante D. Esta aparece en la India en mayo infectando a 320.000 personas al día. Dos meses después en un país de 1500 millones de personas infectaba a 30.000. Tiene que variar toda la proteína de anclaje del virus para que el anticuerpo no lo reconozca y lo elimine. Las farmacéuticas te contarán chino. El negocio es el negocio. La estadística no engaña. A día de hoy la probabilidad de morir por covid es 1 entre 1 millón. 10 veces más que te toque la lotería
La inmuno-oncología, también conocida como inmunoterapia contra el cáncer, es un tratamiento que aprovecha el poder de nuestro sistema inmunológico para combatir el cáncer. La eficacia selectiva de las inmunoterapias continúa impulsando soluciones innovadoras que pueden facilitar un interrogatorio molecular más profundo de la biología del cáncer.
Los investigadores están adoptando cada vez más tecnologías de alto rendimiento que capturan detalles morfológicos, para dilucidar las interacciones adhesivas entre células y matriz y su gobernanza de la arquitectura y función de los tejidos. Al integrar información contextual espacial , pueden mejorar la granularidad de la expresión génica y los hallazgos del análisis de proteínas y generar una descripción más completa del paisaje tumoral altamente complejo y heterogéneo.
A la luz de esto, NanoString desarrolló GeoMx® Digital Spatial Profiler (DSP) para permitir una caracterización multiplexada y resuelta espacialmente de objetivos de ARN y proteínas dentro de secciones de tejido de interés clínico.
La capacidad de la biología espacial para redefinir la precisión en inmuno-oncología se destacó durante un evento GeoCast patrocinado por NanoString que se llevó a cabo el 9 de abril de 2019. Se invitó a oradores invitados a discutir el perfil espacial en inmunoterapia del cáncer y presentar datos utilizando la plataforma GeoMx DSP .
Omer Bayraktar, Ph.D. , del Wellcome Sanger Institute , enfatizó la importancia de preservar el posicionamiento espacial y la heterogeneidad celular dentro de la arquitectura del tejido al cuantificar el transcriptoma. La organización de las células residentes dentro de un tejido influye mucho en su función. Para comprender a fondo la patología de los tejidos y los mecanismos alterados en las enfermedades, por lo tanto, debemos observar la actividad tanto dentro de una célula como entre las células vecinas.
También destacó el valor de analizar simultáneamente múltiples tipos de analitos (ARN y proteínas) y una multitud de objetivos (high-plex). Si bien la función del tejido se refleja típicamente en la firma del ARN subyacente, está dictada directamente por las proteínas. Y la capacidad de multiplexación garantiza un alto nivel de sensibilidad para la creación de perfiles de una sola celda o casi de una sola celda.
Jon Zugazagoitia, MD, Ph.D. , de la Facultad de Medicina de la Universidad de Yale , se refirió al potencial traslacional del perfil espacial de alto plex desde una perspectiva de descubrimiento de biomarcadores clínicos. El perfil espacial se puede utilizar para capturar la heterogeneidad temporal y espacial de la expresión génica entre subgrupos de cáncer.
Al incorporar información espacial, podemos descubrir nuevos biomarcadores que predicen la sensibilidad a la inmunoterapia. Esto se puede utilizar para la estratificación de pacientes en la selección del tratamiento y el diseño de fármacos dirigidos más eficaces. La utilidad clínica del análisis espacial, sin embargo, depende de su aplicabilidad a la práctica clínica, así como de los componentes individuales del generador de perfiles y la capacidad para cuantificar marcadores de distintos compartimentos tumorales.
E. Aubrey Thompson, Ph.D. , de la Clínica Mayo , utilizó un análisis definido espacialmente para estudiar las interacciones inmunitarias del tumor y evaluar su predictibilidad de la respuesta al tratamiento del cáncer. La inclusión de relaciones espaciales célula-célula nos ayuda a extraer características inmunitarias específicas que influyen en gran medida en el resultado del paciente. También se refirió a la necesidad de centrar dicho análisis en una pequeña región de interés para mejorar la sensibilidad de detección.
Sergio Rutella, MD, Ph.D. , del Centro de Investigación del Cáncer John van Geest , exploró la respuesta variable a los tratamientos de inmunoterapia aprovechando la información sobre la topografía del tumor. Hizo un perfil espacial de las biopsias de médula ósea para interrogar la calidad y distribución de las poblaciones de células inmunitarias que se infiltran en el tumor.
Este enfoque lo ayudó a diseccionar aún más el complejo panorama del microambiente tumoral y evaluar la relación entre su contextura inmune y la respuesta al tratamiento. Señaló que la flexibilidad en la personalización de paneles es esencial en su trabajo dada la naturaleza en constante evolución del campo.
Joseph M. Beechem, Ph.D. , del equipo de Investigación y Desarrollo de NanoString , explicó las consideraciones basadas en la investigación que se utilizaron para diseñar el DSP de GeoMx. El generador de perfiles examina tanto el ARN como las proteínas debido a su complementariedad en el descubrimiento de biomarcadores. También utiliza un microscopio adaptativo para adaptarse a diferentes geometrías de tejido.
Cuando se lee un artículo también escrito como este donde se preocupa de las mutaciones que tiene el ADN para convertirse en ARN y en proteína la pregunta es, y esto no se puede decir de forma más sencilla.
Esa forma sencilla empieza por conocer cuál es la alteración que tiene el ADN en el individuo que genera un cáncer posteriormente y como sobre esa alteración agentes externos terminan produciendo un tumor.
Todo esto tiene una lógica aplastante hay que conocer el origen de las cosas para manejarlas.
Primero la alteración de la multiplicación genética, sobre esta alteración previa, producto de la multiplicación casi infinita del ADN y de los errores que se intercalan, un agente externo de los millones que existen produce una metilación de un cromosoma y lo demás es simplemente esperar.
El ARN que se produzca en este núcleo celular va a producir una proteína, que inicia el castigo y conduce al tumor.
Esto está a muchos kilómetros de las ideas primitivas de la genesis de los tumores y de otras muchas patologías.
Primero conocer la mutacion genética existente, y si se puede controlarla.
Después cuando seamos mas sabios, bloquear el agente externo y si no se puede bloquear, modificar el ARN que induce a las proteínas lesivas dentro del citoplasma.
Esto no solo no es nuevo sino conocido a nivel del gran publico aficionado al tema.
La dra. Kariko ha dado un paso en este campo, produciendo un ARN mensajero, que induce una proteína que actúa como antígeno desencadenante de la respuesta inmunitaria. Una Vacuna. La inmunoterapia para el cáncer es un resultado directo de los avances en la tecnología para la generación de anticuerpos recombinantes , la ingeniería de la terapia celular y genética y la creciente apreciación del papel del sistema inmunológico en la prevención y el control del cáncer. Se ha implementado con éxito en la clínica para cánceres hematológicos, como la leucemia linfocítica crónica (CLL), el linfoma no Hodgkin (NHL)y leucemia linfocítica aguda (ALL) . Si bien los tumores sólidos han demostrado ser más difíciles de atacar, la inmunoterapia está ganando terreno para el tratamiento en algunos casos. Una vez considerada un tratamiento de último recurso, la inmunoterapia suele ser la primera línea de tratamiento para el melanoma avanzado y metastásico .
Estamos muy sensibilizados a las malas noticias y nos hacen sufrir sin la menor de las dudas y agregan un componente emocional a los que ya tenemos.
Hace falta estar atento pero no más asustados
La respuesta frente a la nueva variante ha de ser reducir al máximo la exposición al virus y aumentar la población inmunizada en todos los países del planeta, limitando las oportunidades de que aparezcan nuevas mutaciones en el virus.
istockphoto Fernando González Candelas 29/11/2021
La identificación de una nueva variante del SARS-CoV-2 en Sudáfrica, caracterizada por un gran número de mutaciones (55 en todo el genoma, 32 en la proteína S o espícula) y el aumento vertiginoso de su incidencia relativa en esa población ha disparado una vez más las alertas a escala mundial. Varios países han cerrado el tráfico aéreo con Sudáfrica y hay expertos que indican que es “la variante más preocupante que hemos visto hasta la fecha”. La OMS la ha elevado a la categoría de “variante preocupante” y la ha designado con la letra griega “ ómicron”.
Pero, con los datos disponibles, ¿podemos aceptar la pertinencia de estas afirmaciones?, ¿se basan en demostraciones o son conjeturas?, ¿cuándo podemos definir una nueva variante como de preocupación y qué consecuencias tiene eso sobre nuestra estrategia frente a la pandemia? Intentaré en los siguientes párrafos arrojar un poco de luz sobre estos temas.
La secuencia genómica de la variante ómicron (linaje B.1.1.529 en el sistema PANGO, o linaje 21K de NextStrain) muestra 55 mutaciones respecto al virus original de Wuhan, 32 de ellas situadas en la proteína S o espícula, la más importante por su papel en la infección de las células y la respuesta inmunitaria.
Muchas de esas mutaciones se han detectado previamente en variantes de preocupación (VOCs) o de interés (VOIs) del virus, como las mutaciones N501Y (presente en las VOCs alfa, beta y gamma), las T95I, T478K y G142D (todas en delta), o se ha demostrado su papel en la interacción con el receptor celular ACE2 (S477N, Q498R), o se encuentran en regiones de unión de algunos anticuerpos (G339D, S371L, S373P, S375F).
Esta acumulación de mutaciones con efectos conocidos ya es motivo de interés y preocupación, pero todavía se tienen que realizar los experimentos adecuados para demostrar sus efectos cuando se encuentran simultáneamente.
Los efectos de dos mutaciones no son siempre aditivos y las interacciones (epistasias en lenguaje técnico) pueden ser tanto en sentido positivo (aumentando el efecto de cada una) como negativo (disminuyéndolo).
Hasta que no dispongamos de resultados de laboratorio y de datos epidemiológicos y de vigilancia genómica que nos demuestren una mayor transmisibilidad o mayores posibilidades de escape frente a la respuesta inmunitaria no es razonable pasar de vigilancia a alerta o, menos aún, a alarma.
Con la que está cayendo en España con el volcán de la isla de la Palma este artículo es por lo menos estremecedor pero muy curioso de forma pues que me atrevo a copiarlo
Según un estudio de la Universidad de Rutgers (Nueva Jersey, EEUU), las erupciones volcánicas contribuyeron al colapso de las dinastías en China en los últimos 2000 años, al enfriar temporalmente el clima y afectar a la agricultura. La caída de una dinastía se describe a menudo en términos de “colapso”, según el estudio.
Los guerreros de terracota
Está comprobado que las grandes erupciones pueden crear nubes con el potencial de bloquear parte de la luz solar durante uno o dos años. Este tipo de situaciones reducen el calentamiento estival de la tierra en Asia, por lo que habrán monzones más débiles con menos lluvias y, por ende, una reducción en las cosechas. Alan Robock, profesor del Departamento de Ciencias Ambientales en la Escuela de Ciencias ambientales y biológicas afirma: “Hemos confirmado por primera vez que el colapso de las dinastías en China durante los últimos 2000 años son más probables en los años posteriores a las erupciones volcánicas”. Pero existen otras variables que hacen que el estudio sea complejo, “porque si hay guerras y conflictos, las dinastías son más susceptibles al colapso”. Sin embargo nos encontramos con una posible retroalimentación, pues “el impacto de un clima frío en los cultivos puede hacer que los conflictos se den con más probabilidad”.
El equipo de Rutgers ha analizado 156 erupciones volcánicas explosivas desde el año 1 d. C. hasta el año 1915. Ha encontrado niveles elevados de sulfato en los testigos de hielo extraídos de Groenlandia y la Antártida. Pero no se quedaron en el estudio de campo y laboratorio: investigaron la documentación histórica de China sobre 68 dinastías y las guerras ocurridas entre 850 y 1911. Tomaron erupciones volcánicas importantes y vieron en una ventana de aproximación de entre -10 y 2 años si había ocurrido algún colapso. Ocurrió en 62 de los 68 casos. Como bien sabemos, correlación no implica causalidad, sin embargo, tanto datos como causas son, al menos, verosímiles. Mediante el método de Montecarlo generaron erupciones volcánicas aleatorias y, efectivamente, el número promedio de erupciones ocurridas en la realidad en la ventana temporal elegida era superior a la esperada por el azar, con un intervalo de confianza del 99,95 %, mientras que el promedio en ventanas adyacentes era uniformemente menor.
Es realmente complicado demostrar una correlación directa entre erupciones volcánicas y cambios de regímenes gubernamentales, pues la cantidad de variables es enorme. Más cuando hablamos de personas. Incluso con creencias religiosas o sobrenaturales. El “mandato del cielo” fue un concepto muy usado en la filosofía china tradicional que sirvió para mantener la estabilidad entre tanta ida y venida de gobernantes, contribuyendo a que existiese cierta continuidad entre las dinastías. Tanto las élites como los plebeyos aceptaban mejor una cambio de situación si la orden era de origen extraterrenal que si se producía mediante una toma violenta de poder. Algunos de los colapsos ocurrieron con cierta rapidez, en el contexto de un conflicto intenso, con una disrupción socioeconómica significativa y con pérdida de población. Pero otros se dieron como transiciones menos disruptivas, con una continuidad considerable desde el punto de vista burocrático y económico. Estos periodos se sitúan en los “ciclos dinásticos”, en los que hay una época de esplendor, seguida de un declive y un colapso final, atribuido tradicionalmente a la inmoralidad o corrupción de la familia gobernante. Aunque controvertido, las tensiones climáticas han comenzado a tomar un papel importante en el estudio histórico. Los colapsos de las dinastías Tang (907), Yuan (1368) y Ming (1644) están vinculadas a periodos de sequía y frío. El vulcanismo explosivo se propone como un elemento reforzador asociado a colapsos individuales específicos. Los impactos de fío y sequía producidos por las erupciones volcánicas pueden, a su vez, derivar a la muerte del ganado, la degradación acelerada de la tierra y el daño adicional a los cultivos debido a la supervivencia de las plagas agrícolas.
El beneficio del cambio
Volcán en Ulan Hada, Mongolia Interior (China)
Este original trabajo ha sido recientemente publicado en la revista Communications Earth & Environment, bajo el título Volcanic climate impacts can act as ultimate and proximate causes of Chinese dynastic collapse. El hallazgo histórico-científico presentado hace especial énfasis en la necesidad de que exista una buena preparación para futuras erupciones. Especialmente en regiones con poblaciones económicamente vulnerables, como las dinastías Ming y Tang, o con una mala gestión de recursos. Las erupciones de los siglo XX y XXI han sido menores que las históricas en la China Imperial. Sin embargo, las erupciones moderadas también pueden haber afectado de alguna manera a las sequías de Sahel en las décadas de los 70 y 90 del siglo pasado, contribuyendo a la muerte de 250 000 personas y el desplazamiento de diez millones de refugiados. En bastante posible que el cambio climático unido a erupciones importantes afecten dramáticamente a las regiones más pobladas y marginadas del planeta.
Sin embargo, en el artículo no solo se mencionan los efectos directos de las erupciones volcánicas, sino también los ocurridos sobre los ciudadanos. Las dinastías podrían haber aprovechado cualquier tipo de fenómeno natural para ampararse en el “mandato del cielo”. Y lo cierto es que el cambio de gobierno algunas veces han sido beneficiosos, tal como se discute en el propio artículo:
“De hecho, en lugar de ser un fracaso de la sociedad, el colapso en este contexto podría entenderse más como una adaptación a tensiones ambientales, políticas y de otro tipo de situaciones amenazantes, como consecuencia de una concepción cultural (socioecológica) profundamente arraigada a la naturaleza del gobierno, legitimidad dinástica y cambio”.
Me gustaría saber la opinión de Harari sobre este artículo, que parece más producto de la imaginación qué de circunstancias reales.
Pero dada la evolución con qué estamos contemplando en nuestros días los algoritmos, quién se atreve a decir que quién se atreve a decir que no.
Cabe la pregunta de si esto es casualidad o si está ordenado por los dioses. Cierto es que se presenta así y puede ser interpretado como todo en este universo de formas varias.
Algo así como decía mi tío Alfonso siempre pasa lo mismo sucede igual. Pero también se puede decir lo contrario no siempre pasa lo mismo cuando sucede igual.
Hacerse la pregunta de si todo está escrito hoy producto de una evolución que se ríe de los humanos porque todavía no tienen una inteligencia suficiente como para regular esta evolución.
Hara falta otra civilización o otra especie, que apoyándose en los algoritmos, de mas luz a las casualidades
La idea de Harari, de que se acabó, el homo Sapien Sapiens, y nos sucede el de los algoritmos, no me parecen descabelladas. Es posible que este escritor-historiador, fabule a ratos, pero la llegada a gran escala de la civilización digital, ya no permite, ni siquiera la civilización de la razón. El hombre tiene lo suficiente, con lo aportado, por los algoritmos que le proporciona la informática, para tener modelos de como puede proceder, usándolos como modelos referenciales.
Bastantes años ha sufrido los mandamientos impuestos por todas las organizaciones, religiosas y políticas, para tener la posibilidad, basadas en sus propias negligencias, para tomar decisiones, desde su persona, sus necesidades y su información.
Tuvo que venir una potente maquina, que tiene todos tus datos, tus querencias, lo que te hace emocionar, que puede saber todo de ti, para dictarte lo que mas te conviene. La idea que tu sistema referencial, no esta en el medio, sino en big data, que los ordenadores guardan sobre ti en todos y cada uno de tus momentos, sobre ti y tu conducta, puedan dictarte lo que mas te conviene.
Seguro que no va a ser tan fácil, pero lo será mas que los que hemos tenido hasta ahora, donde el medio de los políticos, religiosos y demás mandadores, te hacían la vida no demasiado dulce.
Esto seguro que va a costar su imposición , pero es cuestión de no mucho tiempo.
Lo malo, son siempre lo malvados que se aprovechan de todo para enriquecerse y mandar y enajenarnos. Pero esto no es cuestión de elección, esta ya imponiéndose y a buen paso
Harari parte enesta charla lo descritos en sus libro. Homo Deux, Sapiens y 21 lecciones para el siglo XXI. Lo que diferencia a los humanos, al homo sapiens de otros homos y en general de otras especies de animales, es que los humanos pueden tener una cooperación flexible en grandes cantidades.
Otros animales pueden cooperar tambien, los chimpancés pueden cooperar, pero no a gran escala, grupos de quizás 15, 20, 30, pero ciertamente no en los cientos de miles que son los humanos.
Las abejas pueden operar a gran escala, pero no de manera flexible. Lo que distingue a los humanos es su capacidad para poder operar en estas grandes escalas y operar de manera cooperativa y flexible. Nuestra capacidad para creer en tales cosas es en realidad parte de lo que nos hace tan especiales.
Eso, en última instancia, es la imaginación de la humanidad y nuestra capacidad para creer en las ficciones, para mantener colectivamente una creencia en las ficciones, que es lo que nos da nuestra notable capacidad.
Por lo tanto, nuestras noticias falsas y nuestros hechos alternativos y nuestra voluntad de creer en esto, aunque pueden tener algunas consecuencias graves, también pueden reflejar lo que nos ha traído aquí en primer lugar.
Existen dos aspectos de la realidad.
La realidad objetiva que compartimos con los animales,
y luego está la realidad ficticia, que es lo que nos hace únicos y especiales.
Y las cosas que él incluye en la realidad ficticia, son la mayoría de las cosas que nos son más cercanas y queridas, cosas como religión, corporaciones, naciones, derechos, leyes, países, incluso dinero. Todas estas son ficciones, pero son ficciones tremendamente poderosas. Ellos son los que nos dan el pegamento que nos mantiene unidos y nos permite operar con un poder tan notable.
Según Harari, el poder de la humanidad está en su capacidad de contar y creer en grandes historias.
Hablar hoy no sobre el pasado, no sobre cómo llegamos aquí, sino sobre el futuro, cómo procedemos desde aquí.
Y de hecho, no parece que al Homo sapiens le quede mucho futuro. Probablemente somos una de las últimas generaciones de Homo sapiens, en un siglo o dos como máximo, supongo que los humanos desaparecerán, y la tierra estará dominada por muy diferentes tipos de seres o entidades.
Seres que serán más diferentes de nosotros que de los neandertales o de los chimpancés. Y estamos comenzando a ver los signos de esta revolución a nuestro alrededor.
Un signo particularmente importante, es `preguntar”
Que es lo que le está sucediendo a la autoridad ahora que se está alejando de los humanos. Durante siglos, durante miles de años en realidad, hemos estado acumulando poder y autoridad en nuestras propias manos humanas, pero ahora, la autoridad está empezando a desvanecerse, a pasar de nosotros a otras entidades y en particular, a pasar de nosotros a los algoritmos. Los últimos siglos, vieron el apogeo de la autoridad humana, en la forma de la ideología o cosmovisión conocida. como humanismo.
Durante siglos antes, los humanos creían que la verdadera fuente de autoridad en el mundo, no eran la imaginación, esta no proporcionaba el poder colectivo, la mayoría de los humanos pensaba que la autoridad en realidad viene de afuera, la autoridad viene de arriba de las nubes, de los cielos, la autoridad. viene de los dioses. Las fuentes de autoridad más importantes en política, economía y ética eran los grandes dioses y sus libros sagrados y sus representantes en la Tierra. Los sacerdotes, los rabinos, los chamanes, el califa, los papas, etc.
Luego, en los últimos dos, tres siglos, vimos la revolución humanista, una tremenda revolución política, religiosa y ética, que derribó la autoridad de los cielos a la tierra, a los humanos, y dijo que la fuente más importante de autoridad, la fuente suprema de autoridad, en todos los campos de la vida son los seres humanos, y en particular, son los sentimientos y las elecciones libres de los individuos humanos.
Lo que el humanismo le dijo a los humanos es que cuando tienes algún problema en tu vida, ya sea un problema en tu vida personal, o si es un problema en la vida colectiva de una sociedad entera, o de una nación entera, no necesitas buscar la respuesta en Dios o de la Biblia o de algún hermano mayor. Necesitas buscar la respuesta dentro de ti mismo, en tus sentimientos, en tus elecciones libres.
Y todos hemos escuchado miles de veces estos lemas que nos dicen, escúchate a ti mismo, conéctate contigo mismo y sigue tu corazón, haz lo que te sienta bien. Y estas son realmente las ideas o consejos más importantes sobre la autoridad durante los últimos dos o tres siglos.
En casi todos los campos de la vida, si partimos de la política, es decir , en un mundo humanista, en un mundo que cree que la máxima autoridad son los sentimientos humanos, en la política esto se manifiesta en la idea de que el votante es la autoridad suprema, el votante. sabe mejor. Cuando tienes una decisión política realmente importante, como quién debería gobernar el país, quién debería ser presidente, para que no le preguntes a Dios y no le preguntes al Papa, y no le preguntes al Consejo de Premios Nobel.
En su lugar, vas a todos y cada uno de los Homo sapiens y te preguntas: «¿Qué piensas?
» ¿Qué sientes acerca de esta pregunta? «.
Y la mayoría de las veces, son realmente los sentimientos los que deciden el problema. No es el pensamiento racional , son sentimientos humanos. Y la suposición común en la política humanista es que no hay una autoridad más alta que los sentimientos humanos.
No puedes acercarte a los humanos y decirles: «Sí, piensas así, te sientes así.
«Pero hay alguna autoridad superior » que te dice que estás equivocado «.
Este fue el caso, por ejemplo, en la Edad Media, pero no en la política humanista moderna, ciertamente no en una democracia. Lo mismo ocurre también en el campo económico.
¿Qué es la economía humanista? Es una visión económica que dice que la máxima autoridad son los sentimientos, los deseos, los caprichos de los clientes individuales. El cliente siempre tiene la razón.
¿Cómo saber si una decisión económica es la correcta?
¿Cómo saber si un producto es un buen producto?
En una economía humanista, le preguntas a los clientes, los clientes votan con su tarjeta de crédito, y una vez que han votado, no hay una autoridad superior que pueda decírselo a los clientes «. No, estás equivocado «.
Digamos que quieres producir el mejor automóvil del mundo, ¿cuál es el mejor automóvil del mundo?
Entonces, digamos que Toyota o Ford quieren producir el mejor automóvil del mundo. Reúnen el Nobel Los ganadores del premio de física en química en economía. Ellos reúnen más psicólogos y sociólogos, incluso incluyen a los mejores artistas y los ganadores del Oscar y lo que sea. Y les dan cuatro años para pensar juntos y diseñar y fabricar el coche perfecto.
Y lo hacen. Y producen el Toyota perfecto o el Ford perfecto.
Y luego producen millones de estos automóviles y los envían a agencias de automóviles en todo el país o en todo el mundo.
Y LOS CLIENTES NO COMPRAN LOS COCHES.
Qué significa eso? ¿Significa que los clientes se equivocan? No, el cliente siempre tiene la razón, en una economía humanista. Significa que todas estas personas sabias están equivocadas. No son una autoridad superior al cliente.
En una dictadura comunista, puedes acercarte a la gente y decir: «Este es » el coche para ti
» , decidió el Politburó con su sabiduría,
» que este es el coche perfecto para el trabajador soviético
«y este es el coche que necesitas». vas a tener «.
Pero no es así como funciona en una economía humanista liberal, no hay una autoridad más alta que el cliente. La misma idea también se encuentra en la base del arte moderno.
¿Qué es el arte humanista? Es arte que cree que la belleza está en los ojos del espectador. ¿Cómo sabes qué es el arte? ¿Cómo sabes qué es el buen arte? ¿Qué es hermoso y qué es feo?
Durante miles de años, los filósofos , pensadores y artistas tuvieron todo tipo de teorías sobre qué es arte y qué es bello. Y, por lo general, pensaban que había algún criterio objetivo. Probablemente una vara divina que define el arte y la belleza. Dios define qué es arte y qué es bello.
Luego vino la estética humanista en los últimos dos siglos y cambió la fuente de autoridad a los sentimientos humanos. La belleza esta en los ojos del espectador.
En 1917, hace exactamente un siglo, Que es el bien y que es el mal? ¿Qué es una acción piadosa y qué es un pecado?
Entonces, en épocas anteriores, nuevamente, fue a Dios o fue al Papa, o fue a la Biblia.
Digamos, con el caso de la homosexualidad.
Entonces, en la Edad Media, la Iglesia Católica vino y dijo que la homosexualidad es un pecado. ¿Por qué?
Porque Dios lo dijo, porque la Biblia lo dijo, porque el Papa lo dijo. Y esta fue la fuente de autoridad ética. A nadie le importaba mucho lo que la gente realmente sintiera al respecto.
Ahora, en la era de la ética humanista, el dicho es que no nos importa mucho lo que dice Dios, o lo que dice la Biblia o lo que dice el Papa.
Queremos saber cómo se siente realmente la gente. Si dos hombres están enamorados, y ambos se sienten muy felices con eso, y no hacen daño a nadie, entonces , ¿qué podría estar mal en eso?
Es muy sencillo. La máxima autoridad en el campo de la ética es la autoridad de los sentimientos humanos. Si los humanos se sienten bien por algo y nadie se siente mal por ello, entonces está bien. Por supuesto, también hay algunas cuestiones difíciles en la ética humanista.
¿Qué pasa si me siento bien por algo, digamos una aventura extramarital, pero mi esposo se siente muy mal por eso? Entonces, ¿de quién son los sentimientos más importantes? Entonces tienes un dilema, un dilema ético, tienes una discusión. Pero el punto clave es que la discusión se llevará a cabo en términos de sentimientos humanos, no en términos de mandamientos divinos.
Es interesante que hoy, incluso algunos fundamentalistas religiosos hayan aprendido este truco. Y están usando el lenguaje de los sentimientos humanos y no el lenguaje de los mandamientos divinos.
Vengo de la Universidad Hebrea de Jerusalén, y todos los años, durante los últimos 10 años, tenemos en Jerusalén nuestro propio desfile gay, que es un día muy raro de armonía en Jerusalén, porque esta es una ciudad muy conflictiva. durante la mayor parte del año, pero el desfile gay es un día especial de armonía, porque entonces tienes a todos los judíos ortodoxos, musulmanes y cristianos reunidos para enfurecerse y gritar contra el desfile gay. (audiencia riendo)
Así que tenemos un día de armonía en esta ciudad.
Pero lo realmente interesante son los argumentos que al menos algunos de ellos usan. Van a la televisión o a la radio y dicen que hiere nuestros sentimientos. No dicen que Dios dijo que la homosexualidad es un pecado terrible. Algunos de ellos lo dicen, por supuesto, pero cuando salen a la televisión, generalmente dicen que no deberían realizar el Desfile del Orgullo Gay en Jerusalén, porque hiere nuestros sentimientos.
De la misma manera que los homosexuales quieren que respetemos sus sentimientos, deben respetar nuestros sentimientos y realizar el desfile en Tel Aviv o en algún otro lugar que no sea Jerusalén.
Ahora, de nuevo, realmente no importa lo que piense sobre el argumento, el punto interesante aquí es que incluso esos fundamentalistas religiosos han aprendido que es más efectivo conducir el debate en términos de sentimientos humanos, y no en términos de los mandamientos divinos.
Y finalmente, dado que estamos en una universidad, digamos algunas palabras también sobre educación. Entonces, ¿qué es la educación humanista?
Durante cientos de años, cuando la gente pensaba que la fuente suprema de autoridad estaba fuera de los humanos, el objetivo principal de la educación era conectar a las personas con esa fuente externa de autoridad.
Si pensaba, por ejemplo, que la Biblia era la fuente más alta de autoridad, o que Dios era la fuente más alta de autoridad, entonces el propósito principal de la educación era enseñarle lo que Dios dijo, y lo que la Biblia dijo, y lo que el han dicho los sabios del pasado.
En la educación humanista, dado que la fuente más alta de autoridad son sus propios sentimientos y sus propios pensamientos, el objetivo principal, el objetivo más importante es permitirle pensar por sí mismo.
Vas con todos, desde el jardín de infancia hasta el profesor de esa universidad, y les preguntas: «¿Qué estás tratando de enseñar a los niños » , a los alumnos, a los estudiantes? «.
Y ellos te dirán:» Oh, tratamos de enseñar matemáticas «y tratamos de enseñar física » , y tratamos de enseñar historia» , pero sobre todo, » trato de enseñar a mis alumnos a pensar por sí mismos «.
Este es el ideal más elevado de la educación humanista, porque es la fuente de autoridad, tienes que poder conectarte con ella.
Ahora, todo esto está ahora bajo la nube, ahora enfrenta una gran amenaza, una amenaza mortal.
Toda esta cosmovisión humanista se enfrenta a una amenaza, no tanto de los fundamentalistas religiosos como de los dictadores en Corea del Norte o Rusia o en cualquier otro lugar.
La gran amenaza para la cosmovisión humanista ahora está emergiendo de los laboratorios, de las universidades, de los departamentos de investigación en lugares como Silicon Valley, porque lo que más y más científicos nos dicen es que esto toda la historia del humanismo se basa en realidad de la ciencia o una comprensión anticuada del mundo y , en particular, una comprensión anticuada del Homo sapiens, de este simio de África Oriental.
El humanismo se basa en la creencia en el libre albedrío, en la capacidad de los seres humanos para tomar decisiones libres y en una creencia muy fuerte en los sentimientos humanos como la mejor fuente de autoridad en el mundo.
Pero ahora los científicos están diciendo, en
primer lugar, que no existe el libre albedrío.
La ciencia está familiarizada con solo dos tipos
de procesos en la naturaleza.
Tienes procesos deterministas y tienes procesos aleatorios y ,
por supuesto, tienes combinaciones de aleatoriedad
y determinismo, que dan como resultado un resultado probabilístico, pero nada de eso es libertad.
La libertad no tiene absolutamente ningún significado desde una perspectiva física o biológica.
Es solo otro mito, otro término vacío que los humanos han inventado.
Los humanos han inventado a Dios,
y los humanos han inventado el cielo y el infierno,
y los humanos han inventado el libre albedrío,
pero no hay más verdad para el libre albedrío
que el cielo y el infierno.
Y en cuanto a los sentimientos, definitivamente son reales,
no son una ficción de nuestra imaginación,
pero los sentimientos son en realidad solo algoritmos bioquímicos, y no tienen nada de metafísico
o sobrenatural.
No hay ninguna razón obvia para considerarlos
como la máxima autoridad del mundo y, lo que es más importante, lo que los científicos e ingenieros nos dicen cada vez más es que si solo tenemos suficientes datos y suficiente poder de cómputo, podemos crear algoritmos externos. que comprenden a los humanos y sus sentimientos mucho mejor de lo que los humanos pueden comprenderse a sí mismos.
Y una vez que tienes un algoritmo que te comprende
y comprende tus sentimientos mejor de lo que te entiendes a ti mismo, este es el punto en el que la autoridad realmente se desplaza de los humanos a los algoritmos.
Eso suena a la vez aterrador, supongo, y complicado.
Así que intentaré explicar qué significa.
En primer lugar, ¿qué significa que los sentimientos y los organismos son realmente algoritmos?
Se puede decir que, en cierto modo, es posible resumir más de un siglo de investigación en las ciencias de la vida y especialmente en biología y en la evolución, en tan solo tres palabras, los organismos son algoritmos. Esta es ahora cada vez más la visión dominante, no solo en las ciencias de la vida, sino también en la informática, razón por la cual las dos se están fusionando,
y lo que esta idea de que los organismos son algoritmos,
lo que realmente significa es que los sentimientos humanos
y no sentimientos, sensaciones y emociones sólo humanos, también de chimpancés, elefantes y delfines, son sólo un proceso bioquímico de cálculo, calculando probabilidades para tomar decisiones. Los sentimientos no son una cualidad metafísica
que Dios le dio al Homo sapiens para escribir poesía
y apreciar la música, los sentimientos son procesos de cálculo,
cálculo bioquímico, moldeados por millones de años de selección natural, para permitir a los humanos y otros mamíferos
y otros animales hacer el bien. Decisiones cuando se enfrentan a problemas de supervivencia y reproducción.
Qué significa eso?
Tomemos un ejemplo concreto.
Digamos que eres un babuino en algún lugar de la sabana africana y te enfrentas a un problema típico de supervivencia. Para sobrevivir, tienes que comer, para sobrevivir tienes que tener cuidado de no ser comido por otra persona. Y digamos que mientras caminas por la sabana, de repente ves un árbol con plátanos. Pero también ves un león, no lejos del árbol. Y debe tomar una decisión sobre si arriesgar su vida por los plátanos o no.
Este es el tipo de problema que los animales como los babuinos y los humanos se han enfrentado durante millones y millones de años. Ahora para esto es realmente una cuestión de calcular probabilidades. Yo, el babuino, necesito calcular la probabilidad de que me muera de hambre si no como los plátanos, frente a la probabilidad de que el león me coma si trato de alcanzar estos plátanos. Necesito saber qué probabilidad es mayor para tomar una buena decisión.
Para eso, primero que nada necesito recopilar muchos datos, necesito datos sobre los plátanos. ¿Qué tan lejos están los plátanos? ¿Cuántos plátanos? ¿Son grandes o pequeños? ¿Maduro o verde?
Es una situación cuando hablamos de 10 plátanos grandes maduros, y es muy diferente si solo tienes dos plátanos verdes pequeños. Del mismo modo, necesito información, necesito datos sobre el león.
¿Qué tan lejos está el león? ¿Qué tan grande es el león? Qué rápido creo que puede correr el león. ¿El león está dormido o despierto? ¿El león parece hambriento o saciado? Necesito todo este tipo de datos sobre el león. Y, por supuesto, necesito muchos datos sobre mí, Como estoy de hambriento tengo, qué tan rápido puedo correr, etc. Entonces necesito tomar todos estos datos y de alguna manera calcularlos muy, muy rápido, las probabilidades. ¿Cómo lo hace el babuino? El babuino no saca un bolígrafo y una hoja de papel o calculadora y comienza a calcular probabilidades. No, todo el cuerpo del babuino es la calculadora. Toma los datos con lo que llamamos nuestros sentidos, nuestras sensaciones, con nuestra vista, con nuestro olfato, con nuestros oídos, tomamos todos los datos también desde dentro del cuerpo, y luego del sistema nervioso y el cerebro, ellos ‘ Si la calculadora, en una fracción de segundo, calcula las probabilidades, y la respuesta no aparecerá como un número, la respuesta aparecerá como un sentimiento o una emoción.
Si hay posibilidades de que vaya a buscar los plátanos, entonces esto aparecerá como la emoción del coraje. Me sentiré muy valiente, siento que puedo hacerlo, o mi cabello se destacará, mi pecho se hinchará y correré por los plátanos. Si el resultado del cálculo es que es demasiado peligroso, entonces esto también aparecerá no como un número, sino como una emoción, esto es miedo. Así es como surge el miedo. Y estaré muy asustado y huiré de allí. Entonces, lo que en el lenguaje cotidiano llamamos sentimientos y emociones y demás, de acuerdo con la teoría estándar actual en las ciencias de la vida, estos son en realidad algoritmos bioquímicos que calculan probabilidades.
Ahora hasta hoy, hasta principios del siglo XXI, esta idea, esta comprensión de que los organismos son algoritmos, que las emociones y los sentimientos son en realidad solo un proceso bioquímico de cálculo de probabilidades, esto no tuvo mucho impacto práctico, porque nadie tenía la capacidad para recopilar suficientes datos, y nadie tenía la capacidad informática necesaria para analizar esos datos y comprender realmente lo que está sucediendo dentro de mí.
Durante miles de años, todo tipo de autoridades intentaron hackear a los humanos, intentaron comprender lo que pasa por nuestra mente, ¿qué pensamos? Que sentimos
Pero nadie pudo hacerlo realmente.
La Iglesia Católica en la Edad Media o la KGB en la Rusia Soviética, estaban muy interesados en entender a los humanos, en piratear humanos, pero no pudieron hacerlo. Incluso si la KGB te seguía por todas partes y registraba cada conversación que tenías y cada actividad que realizabas , la KGB no tenía el conocimiento biológico y no tenía la capacidad de computación necesaria para realmente entender lo que sucede dentro del ser humano. cerebro o dentro del cuerpo humano.
Así que hasta hoy, cuando el humanismo le dijo a la gente, no escuches al Papa, no escuches la Biblia, no escuches a Stalin, no escuches a la KGB, escuches tus sentimientos.
Este fue un buen consejo.
Debido a que sus sentimientos realmente fueron los mejores métodos para tomar decisiones, son el mejor algoritmo del mundo.
Tus sentimientos, los sentimientos humanos, fueron algoritmos moldeados por millones de años de selección natural, algoritmos que resistieron las pruebas de calidad más duras del mundo, las pruebas de calidad de la selección natural. Entonces, si tuviste que elegir entre escuchar la Biblia y escuchar tus sentimientos, fue un muy buen consejo, lo que te dijo el humanismo, escucha tus sentimientos. La Biblia era solo la sabiduría de algunos sacerdotes en la antigua Jerusalén.
Tus sentimientos fueron el mejor mecanismo para tomar decisiones que ha sido moldeado por millones y millones de años de selección natural. Así que era un buen consejo escuchar tus sentimientos. Pero ahora las cosas están cambiando. Ahora estamos en la intersección o en el punto de colisión entre dos inmensos maremotos científicos. Por un lado, durante más de un siglo, al menos desde Charles Darwin, hemos ido adquiriendo una comprensión cada vez mejor del cuerpo humano, y del cerebro humano, y de la toma de decisiones y las emociones y sensaciones humanas, etc. Y al mismo tiempo, con el auge y el desarrollo de las ciencias de la computación, hemos estado aprendiendo a diseñar cada vez mejores algoritmos electrónicos, con cada vez más potencia de cálculo.
Hasta hoy, estos dos eran desarrollos separados, pero ahora se están uniendo. Creo que lo crucial que está sucediendo ahora mismo en la segunda década del siglo XXI es la fusión de estos dos maremotos. El muro que separa la biotecnología de la infotecnología se está derrumbando, lo que también se puede ver en el mercado, que corporaciones como Apple o Amazon o Google o Facebook, que comenzaron como corporaciones estrictamente infotecnológicas, se están convirtiendo cada vez más en biotecnología. Porque realmente, ya no hay ninguna diferencia esencial entre los dos. Y estamos muy cerca del punto en que, a diferencia de la KGB, y a diferencia de la Iglesia católica, Facebook o Google podrán entenderte mejor que tú mismo.
Porque ellos tendrán los datos, tendrán el conocimiento biológico y tendrán el poder de cómputo necesario para comprender exactamente cómo se siente y por qué se siente de la manera en que se siente. Esto ya está sucediendo en un campo muy importante que es el campo de la medicina.En el campo de la medicina, ya ha ocurrido básicamente.
La autoridad ya ha comenzado a cambiar drásticamente de los humanos a los algoritmos. Creo que es una estimación o predicción muy justa, que las decisiones más importantes sobre su salud, sobre su cuerpo, durante su vida, no las tomará usted sobre la base de sus sentimientos, sino que serán tomadas por algoritmos sobre la base de lo que ellos saben de ti y tú no te conoces a ti mismo.
Para dar un ejemplo práctico de la vida real, que apareció en muchos titulares. Hace dos, tres años, hubo una historia muy famosa sobre Angelina Jolie. Hizo una prueba genética, una prueba de ADN que reveló que tenía una mutación, creo que estaba en el gen BRCA1. Y según las estadísticas de big data, las mujeres que tienen esta mutación en particular en este gen en particular, tienen un 87% de probabilidades de desarrollar cáncer de mama.
Ahora, en ese momento, Angelina Jolie no tenía cáncer de mama. Por supuesto, hizo todas las pruebas y controles y no tenía cáncer de mama en ese momento. También se sentía perfectamente sana. Sus sentimientos le decían: «Estás perfectamente bien » . No necesitas hacer nada «.
Pero los algoritmos de big data, le estaban contando una historia muy diferente. Le dijeron que tienes una bomba de tiempo en tu ADN. Y aunque no sientes que nada está mal, es mejor que hagas algo al respecto ahora. Y Angelina Jolie, con mucha valentía y sensatez, prefirió escuchar el algoritmo y no sus propios sentimientos. Se sometió a una doble mastectomía, y también publicó su historia, creo que fue en The New York Times con el fin de alentar a otras mujeres a hacer pruebas similares y tal vez tomar medidas preventivas similares.
Entonces, este tipo de escenario, que tus sentimientos te dicen que estás absolutamente bien, pero algún algoritmo de big data que te conoce, te dice que no, que no estás bien y que prefieres escuchar el algoritmo, creo que esto va a ser cada vez más la forma de la medicina en el siglo XXI.
Pero ganó ‘ permanecer restringido a la medicina.
Es probable que veamos un cambio similar en la autoridad y
en casi todos los campos de la actividad humana.
En el pasado, digamos en la Edad Media, tenías las religiones monoteístas y politeístas, diciéndole a la gente que la autoridad viene de las nubes, de los dioses.
Si tiene un problema en su vida, escuche la Palabra de Dios, escuche la Biblia.
Luego estaba el humanismo, viniendo y diciéndole a la gente
que bajemos la autoridad de las nubes a los sentimientos humanos.
No escuches la Biblia, no escuches a Dios, al Papa,
escuches tus propios sentimientos.
Ahora, estamos viendo el surgimiento de una nueva visión del mundo, o una nueva ideología, que podemos llamar dataismo,
porque cree que la autoridad al final proviene de los datos.
Y el dataismo devuelve las autoridades a las nubes,
a Google Cloud, a Microsoft Cloud.
Y el dataismo le dice a la gente: «No escuche sus sentimientos
» , escuche a Google, escuche a Amazon.
«Ellos saben cómo te sientes » y también saben por qué te sientes de la manera en que te sientes «y, por lo tanto, pueden tomar mejores decisiones » en tu nombre «.
¿Qué significa en la práctica?
Es muy importante para mí como historiador, para siempre bajar no solo la autoridad de las nubes a los humanos, sino también llevar estas ideas muy abstractas a ejemplos concretos de la vida diaria de las personas.
Entonces, ¿qué significa, que la autoridad se aleja
de nuestros sentimientos hacia estos algoritmos externos?
Entonces, como escribo libros, me interesan mucho los libros.
Tomemos un ejemplo del mundo de los libros.
¿ Una decisión mundana de que debes tomar qué libro leer?
Entonces, en la Edad Media, fuiste a la fuente de autoridad, al sacerdote, y le preguntaste al sacerdote qué libro debería leer
y el sacerdote te dirá que leas la Biblia, es el mejor libro del mundo, todas las respuestas están ahí, no necesitas leer ningún otro libro en tu vida, solo la Biblia es suficiente.
Luego vino el humanismo en los últimos dos, tres centurias,
y le dijo a la gente, «Ah, sí, la Biblia » ,hay algunos capítulos agradables allí, «pero tantas partes de ella, » realmente necesitan un buen editor.
«Y realmente, no necesitas escuchar a nadie » , excepto a ti mismo, «solo sigue tus sentimientos».
Hay tantos buenos libros en el mundo.
Vas a una librería, deambulas entre los pasillos,
tomas este libro y ese libro, lo hojeas, sientes un instinto
que te conecta con un libro en particular, tomas este libro, lo
compras, lo lees.
Así que esta era la forma humanista de elegir qué libro comprar.
Ahora, con el comienzo del auge del dataismo, vamos a la librería virtual de Amazon, y en el momento en que entro a la librería de Amazon, aparece un algoritmo que me dice: «Te conozco » .
Te he estado siguiendo, » He estado siguiendo tus gustos y disgustos «y, según lo que sé sobre ti » , y las estadísticas sobre millones de otros lectores y libros, «te recomiendo estos tres libros».
Pero este es solo el primer pequeño paso .
El siguiente paso, que ya se está tomando hoy,
es que Amazon con el fin de mejorar el algoritmo,
necesita más y más datos sobre usted, y tus preferencias y tus sentimientos.
Así que si lees, como yo un libro en Kindle, debes saber que mientras lees el libro, por primera vez en la historia, el libro te está leyendo.
Nunca había sucedido antes.
Te remontas a la antigüedad o te remontas a la revolución de la impresión moderna temprana , así que Gutenberg trajo la impresión a Europa y empezaron a imprimir libros, pero todos estos libros impresos por Gutenberg y sus sucesores, nunca leen a la gente.
Son solo las personas que leen los libros. Ahora los libros leen a la gente. Mientras yo o usted leemos un libro en Kindle, Kindle me está siguiendo. Kindle sabe qué páginas leo rápido, que paginas leo lento, para Kindle es muy fácil saberlo. Y Kindle también se da cuenta cuando dejo de leer el libro, y quizás nunca volver a él.
Basado en ese tipo de información, Amazon tiene una idea mucho mejor de lo que me gusta y lo que no me gusta. Pero esto sigue siendo muy, muy primitivo. Lo crucial es llegar a los datos biométricos, meterse dentro de tu cuerpo, para entrar en tu cerebro en tu mente. Y esto puede empezar a suceder hoy. Si conecta el Kindle al software de reconocimiento facial, que ya existe, entonces Kindle podría saber, cuando me río, cuando lloro, cuando estoy aburrido, cuando estoy enojado, según mi expresión facial. Es de la misma manera que sé cuál es tu estado de ánimo ahora cuál es tu emoción al tratar de leer tus rostros.
Ahora podemos hacer que Kindle lea las caras de los lectores. Y en base a eso, Amazon tendrá una idea mucho mejor de lo que me gusta o no me gusta. Pero esto sigue siendo primitivo porque todavía no entra en el cuerpo. El paso realmente crucial,y estamos muy, muy cerca de ese paso, es cuando conectas el Kindle a sensores biométricos en o dentro de mi cuerpo. Y una vez que haces eso, Kindle sabrá el impacto emocional exacto de cada frase que leo en el libro. Leo una frase y Kindle sabe lo que significa que Amazon sabe, ¿Qué pasó con mi presión arterial? que paso con mi nivel de adrenalina, qué pasó con mi actividad cerebral mientras leía esta oración. Para cuando termino el libro, digamos que leí «Guerra y paz» de Tolstoi. Cuando termino de leer «Guerra y paz» de Tolstoi, Olvidé la mayor parte. Pero Amazon nunca olvidará nada. Cuando termine «Guerra y paz», Amazon sabe exactamente quién soy cual es mi tipo de personalidad, y cómo presionar mis botones emocionales. Y basado en este tipo de conocimiento, puede hacer mucho más que recomendarme libros. En diferentes situaciones, podría tomarse de diferente es formas. Si vives, digamos en Corea del Norte, para que todos tengan que usar esta pulser a biométrica, y si entras en una habitación y ves una foto de Kim Jong-un, y su presión arterial y actividad cerebral indican ira, entonces ese es tu fin. En una sociedad liberal, como Estados Unidos, puede ir en diferentes direcciones.
Amazon recomienda no solo cosas como libros, pero cosas mucho más importantes como citas o incluso matrimonios. Digamos que estoy en una relación y mi novio me pregunta, bueno, me quiero casar, o nos casamos o nos separamos, tú tienes una opción. Así que una de las opciones más importantes que cualquier animal necesita hacer durante su vida sobre compañeros. Digamos que se le presenta este dilema, o nos casamos, o nos estamos separando, no hay una tercera opción.
Entonces, en la Edad Media, acudirías al cura para pedirle consejo.
En la era humanista, en los siglos XIX y XX, te dirán que sigas a tu corazón.
Intenta conectarte con tu yo auténtico y solo sigue tu corazón.
In the 21st century, the advice will be «Oh, escucha a Amazon, «pregúntale a Google qué hacer». Y yo venía a Google o Amazon, y yo decía: «Google, tengo este dilema. «¿Qué me recomiendas que haga? «¿Cuál es el mejor curso de acción para mí?». Y Google dirá: «Oh, te he estado siguiendo. «durante tu vida. «He estado leyendo todos tus correos electrónicos «y todas tus palabras de búsqueda en Internet. «He estado escuchando todas tus llamadas telefónicas, «He estado rastreando cada vez que veías una película o leías un libro, «Vi lo que le estaba sucediendo a tu corazón. «Además, cada vez que ibas a una cita, «Te seguí por ahí «y vi lo que le pasó a tu presión arterial, «o a tu ritmo cardíaco o a tu actividad cerebral. «Y conozco tu escaneo de ADN «y todos sus registros médicos y luego lo que sea. «Y, por supuesto, sé que tengo todos estos datos, «también sobre tu novio o tu novia, «y tengo datos sobre millones y millones de éxitos «y relaciones infructuosas. «Y en base a todos estos datos, «Te lo recomiendo con una probabilidad del 87%, «que es una buena idea casarse. «Eso es lo mejor para ti, casarte».
Pero Google me dirá: «Te conozco tan bien, «que incluso sé que no te gusta el consejo que te di ahora. «Y también entiendo por qué, «porque crees que puedes hacerlo mejor, «porque piensas que, oh, no es lo suficientemente guapo».
«Y su algoritmo bioquímico que se formó en: «la sabana africana hace 100.000 años, «le da demasiada importancia «a miradas externas, a la belleza.
«Porque esto es lo que importaba «la sabana africana hace 100.000 años, «Fue un buen indicador de la forma física. «Y todavía estás siguiendo este algoritmo anticuado «de la sabana africana. «Pero mi algoritmo es mucho, mucho mejor.
«Tengo las últimas estadísticas sobre relaciones «en el mundo posmoderno. «Y no ignoro la belleza, «No ignoro la buena apariencia. «Pero a diferencia de tu algoritmo, «que da buen aspecto al 30% en peso, «al evaluar las relaciones, «Sé que el verdadero impacto de la apariencia «en el éxito a largo plazo de una relación es solo del 11%. «Así que tomé esto en consideración, «y todavía te estoy diciendo, «que es mejor que te cases».
Y al final, es una pregunta empírica. Google no tiene que ser perfecto, solo tiene que ser mejor que el humano promedio. Solo tiene que hacer mejores recomendaciones consistentes. que las decisiones que las personas toman por sí mismas. Y la gente a menudo comete errores tan terribles. en las decisiones más importantes de su vida. E ligen qué estudiar o con quién casarse o lo que sea, y después de 10 años, ellos, «Ah, este fue un error tan terrible». Y Google solo tendrá que ser mejor que eso. Y no sucederá de la noche a la mañana, como en alguna revolución inmediata, es un proceso. Le preguntaremos a Google o Amazon o Facebook o lo que sea, por su asesoramiento en más y más decisiones, y si vemos que efectivamente obtenemos buenas recomendaciones mejor
que las decisiones que nosotros mismos solemos tomar,
entonces confiamos cada vez más en ellos, confíe más en ellos con nuestros datos, y también confiar más en ellos con autoridad para tomar decisiones por nosotros. Y realmente está sucediendo a nuestro alrededor en pequeños pasos de bebé.. Como si pensaras en orientarte por la ciudad. Entonces cada vez más personas delegan esta responsabilidad a Google Maps o Waze, o alguna otra aplicación de GPS. Llegas a una intersección tu instinto te dice que gires a la derecha Google Maps te dice: «No, no, no, gira a la izquierda. «Sé que hay un atasco a la derecha». Y confías en tu intuición y doblas a la derecha y llegas tarde. La próxima vez, dices: «Está bien, probaré Google Maps», y sigues la recomendación de Google y llegas a tiempo, aprendes a no confiar en tu intuición, es mejor confiar en Google. Y muy pronto, la gente llega a un punto, cuando no tienen idea de dónde están, pierden su capacidad espacial para saber dónde están y encontrar su camino por el espacio. Simplemente siguen ciegamente cualquiera que sea la aplicación les está diciendo. Y si pasa algo y el teléfono inteligente se apaga, no tienen ni idea. No saben cómo moverse por el espacio.
Esto ya está sucediendo hoy, y no es porque algún gobierno nos obligó a hacerlo, son decisiones que todos tomamos a diario.
Ahora, dos advertencias antes de que abra la palabra para sus preguntas.
En primer lugar, es muy importante enfatizar que todo esto depende de la idea que estamos en proceso de hackear al ser humano, y en particular, de hackear la mente humana y el cerebro humano.
Pero todavía estamos muy, muy lejos de comprender el cerebro e incluso más lejos de comprender la mente. Y existe la posibilidad de que al final, resultará que, después de todo, los organismos no son algoritmos.
Hay algunas cosas muy profundas que todavía no entendemos sobre el cerebro y la mente, y todo este sueño se convertirá en una fantasía. Entonces, esto es lo primero que debemos tener en cuenta, todavía, todavía no estamos allí, comprender realmente el cerebro humano y la mente humana.
La otra cosa importante a tener en cuenta, es que la tecnología nunca es determinista. Puedes usar la misma tecnología con el fin de crear tipos de sociedades muy diferentes. Ésta es una de las principales lecciones del siglo XX.
Podrías usar la tecnología del siglo XX, de la Revolución Industrial, trenes, electricidad, radio, coches, etc. Podrías usar eso para crear una dictadura comunista o un régimen fascista o una democracia liberal. Los trenes no te dijeron qué hacer con ellos.
Aquí tienes una imagen muy famosa. Esto es Asia Oriental. Una imagen del este de Asia por la noche tomada desde el espacio ultraterrestre desde un satélite.
Y lo que ves en la esquina inferior derecha es Corea del Sur, un mar de luz. Y en la esquina superior ves China, otro mar de luz. Y en medio de la parte oscura, no es el mar esto es Corea del Norte. Ahora, ¿por qué Corea del Norte es tan oscura? mientras que Corea del Sur está tan llena de luz? No es porque los norcoreanos no hayan escuchado sobre electricidad o bombillas. Es porque eligieron hacer cosas muy diferentes. con electricidad que la gente de Corea del Sur o China.
Así que esta es una ilustración muy visual de la idea. que la tecnología no es determinista. Y si no le gustan algunos de los escenarios, algunas de las posibilidades que he delineado en esta charla esta noche, debe saber que todavía puede hacer algo al respecto.
La disbiosis está de moda desde hace años y la preocupación por ella es que motiva las enfermedades degenerativas que son el pesar de nuestro tiempo.
La exposición a metales pesados tiene una repercusión directa y provoca una disbiosis aunque se habla mucho de ella, tenemos pocas herramientas para combatirla, porque lo importante no son las bacterias mutadas o alteradas sino la polución que introduce en nuestra biología metales nocivos que altera los gérmenes de nuestras cavidades y pone en marcha la disbiosis.
Es impensable cómo reparar la polución que de una manera universal nos afecta, pero ya tenemos una importante parte del conocimiento.
Las innumerables variedades de enfermedades degenerativas, pueden hechar causante pueden estar causadas por este desarreglo de nuestra biología como consecuencia de la polución.
Quién convence al mundo industrial para que cambie los parámetros de la obtención de la energía.
Pero por lo menos ya tenemos una idea bastante acertada de este grupo terrible de enfermedades pero mutilan y sobre los que solo sabemos poner el parches.
No está de más revisar el tema:
La microbiota intestinal tiene un efecto directo en cómo se gestione esa carga corporal total
La microbiota intestinal afecta a la absorción y metabolismo de metales pesados
A su vez los metales pesados alteran la composición de la microbiota intestinal y producen una disbiosis y como consecuencia van a alterar los perfiles metabólicos.
La disbiosis intestinal inducida por metales pesados puede causar enfermedades metabólicas y esto implica a cualquier tipo de patología y por otro lado que los probióticos pueden ser una buena estrategia para proteger la microbiota intestinal de la exposición a metales pesados ya que pueden cambiar la expresión de proteínas transportadoras de metales y mantener la función tan importante de barrera intestinal.
Los metales pesados contribuyen al desarrollo de enfermedades metabólicas y la repercusión de los metales pesados en enfermedades metabólicas es por la alteración que éstos producen en la microbiota intestinal y esta microbiota alterada es la que produce la mayor parte de las alteraciones metabólicas l
La microbiota intestinal es la primera línea de defensa frente a cualquier tóxico y por supuesto frente a los efectos tóxicos de los metales pesados y hay una relación bidireccional entre ambos.
La exposición a metales pesados altera la composición función y perfil metabólico de la microbiota intestinal y a su vez la vitalidad intestinal altera la absorción y el metabolismo de los metales pesados.
Si hay mucha carga por metales pesados se va a provocar una alteración en la composición de la microbiota intestinal .
Esta alteración de la microbiota intestinal va a hacer perder capacidad
detoxificación y a alterar la absorción de estos metales pesados y alterar la barrera intestinal con lo cual vamos a tener una vía libre desde los metales pesados a la sangre es decir en primera instancia los metales pesados pueden provocar una desvíos una mayor absorción y una peor gestión y degradación de los metales pesados .
Es necesario una detoxificación una descarga de toda esa cantidad de tóxicos para poder revertir la situación intestinal y otras veces si el desorden no es demasiado grande o no se ha sostenido suficiente tiempo trabajando
sobre microbiota intestinal se produce una desintoxicación y una buena gestión
de los metales pesados y de otros tóxicos a los que estemos expuestos
la microbiota intestinal puede afectar a la integridad de la barrera intestinal.
Los desvíos de la microbiota cursan con un aumento de la permeabilidad intestinal que a su vez va a tener más absorción de metales pesados y más absorción de todo tipo de tóxicos que van a llegar a la sangre y a toda nuestra biología.
Esto marca muy bien la relación bidireccional entre los metales pesados y la microbiota intestinal es decir la exposición a metales pesados va a alterar va a afectar a la composición de la microbiota intestinal y va a alterar todo el perfil metabólico de la microbiota .
La microbiota en buenas condiciones va a poder reducir la absorción de metales pesados y también va a hacer la o permite la posibilidad
de conversión de los metales pesados a formas menos tóxicas al hacerlos más solubles.
La alteración de la microbiota intestinal de la desvió sis inducida por metales pesados incrementa todo tipo de alteraciones metabólicas y procesos
alérgicos en general.
Esto se puede extrapolar a cualquier tipo de patología neurodegenerativas y
El uso de probióticos va a odificar la estructura y la función de la microbiota va a promover la producción de ácidos grasos de cadena corta y va a cuidar la integridad y mantener la integridad de barrera del intestino.
La disbiosis que provocan los metales pesados metales pesados cobran interés en los últimos 50 años cuando estamos expuestos de una
manera acumulativa cada vez más hay más exposición por encima es
acumulable el problema de los metales pesados es que no son biodegradables y entonces persisten en el medio ambiente durante muchísimo tiempo. Los mentales pesados que pueden tener una vida media de 25 años con lo
cual ya no es la exposición actual sino toda la exposición acumulada.
Los más importantes respecto a la salud aunque todos son relevantes; son
el mercurio el plomo el cadmio el arsénico el níquel sería más correcto decir
metales tóxicos que metales pesados y así incluimos arsénico y aluminio que
tienen una relevancia desde el punto de vista psicológico muy alta
El mercurio está presente en todas las enfermedades crónicas degenerativas en enfermedades graves en todas aquellas que afectan al sistema nervioso y
La exposición a la que estamos expuestos por todas las vías desde la inhalación la ingestión a estos metales es enorme.
dependiendo de la forma
El mercurio de una manera metálica inorgánica o el metilmercurio que es el más tóxico ( es el que viene en los pescados en los mariscos y que tomamos por ingestión, también tenemos una exposición muy alta mercurio con las amalgamas)
En conclusión la exposición a metales pesados tiene una repercusión directa y provoca una disbiosis
La microbiota intestinal tiene un efecto directo en cómo se gestione esa carga corporal total
La microbiota intestinal afecta a la absorción y metabolismo de metales pesados
El psiquiatra Británico Edward Bullmore en su libro The inflamed mind (la mente inflamada) apunta que la depresión es en realidad un problema inflamatorio
Edward Bullmore, profesor de psiquiatría en la Universidad de Cambridge y director del Wolfson Brain Imaging Center en el Departamento de Neurociencia Clínica.
Esencialmente, ‘The Inflamed Mind’ de Bullmore sienta las bases para el argumento de que parte de la depresión es causada por una inflamación corporal que puede penetrar la barrera hematoencefálica para inflamar las células y redes cerebrales y, por lo tanto, causar distimias de las que predomina la depresión.
La inflamación corporal es el mecanismo universal que utilizan los seres vivos para repararse tras un trauma o tras las erosiones que nuestra biología se produce por vivir. De forma que su origen es múltiple y cada vez que nos ponemos en contacto con mas polución física y psíquica, la inflamación progresa y los desordenes que se sufre como consecuencia de su exceso y persistencia van a hacernos enfermar.
La inflamación es el mecanismo de reparación el estrés es la clínica que produce este mecanismo y la depresión es una de estas manifestación, de absoluta inespecificidad en su aparición.
La inflamación llena la etiología de las enfermedades neurodegenerativas
Por supuesto que interviene el sistema inmunitario, no hay otro, el problema es que en biología todo debe ser ajustado y tanto el defecto como el exceso pueden producir un exceso o un defecto en la inflamación y también un exceso o defecto de sus manifestaciones psíquicas.
Bullmore comienza su argumento presentando la teoría subyacente del libro que incorpora el funcionamiento del sistema inmunológico. Afirma que esta causa de depresión relativamente simple y a simple vista puede haberse pasado por alto a lo largo de los años
Este concepto de unión de lo físico con lo psiquico, lo ha sabido el hombre desde que alboreo.
Uno de sus problemas es de nomenclatura etiológica.
Depresion es una manifestación psíquica, su origen plural es la consecuencia de la alteración de alguna región o de varias del sistema nervioso. De manera rutinaria y después de los trabajos de Seyle, se llama estrés a las manifestaciones clínicas de un trauma.
El sufrimiento físico y ese otro mal llamado psíquico se deben al compromiso de estructuras preferentemente del diencéfalo.
Se sufre por cualquier cosa, aunque a veces se presenta como dominante y excepcionalmente de causa concreta
La parcelacion de las manifestaciones del sistema nerviosas nunca ha sido real y si un intento de entenderlas.
Un dolor de cabeza por una meningitis, nunca se acompaña de alegría y siempre de tristeza
Bullmore postula que la depresión puede haber sido seleccionada para proteger a los antepasados enfermos de las obligaciones sociales y la competencia cuando necesitan descansar o como una forma de cuarentena para las personas infectadas para evitar que la infección acabe con toda una tribu.
El reciente trabajo de que la estimulación de la amigdala del hipocampo, rescata a una enferma con depresión y múltiples intentos de suicidio, intensifica la unión de lo físico con lo psiquico
Inflamación y depresión están vinculados y lo están siempre, es cuestión de que predomina.
Si un dolor de apendicitis pone triste a un enfermo nadie duda de la simbiosis. Si solo aparece depresión acudimos a lo etéreo, a lo social, y nos apartamos de lo orgánico, lo químico que promueve la disfuncion
Pero la estructura nerviosa que proporciona clínica física y psíquica es la misma, posiblemente el lóbulo limbico, de preferencia, aunque su engarce con el lóbulo frontal, le presta cierta modulación.
Bullmore ofrece algunas pruebas para respaldar sus afirmaciones, pero después de una emoción inicial, el libro me deja decepcionado. Algunas teorías interesantes que, si se prueban correctas, pueden ayudar a un subgrupo de personas deprimidas.
Lo importante no es la depresión sino como evitar la inflamación crónica imnecesaria no racional responsable de tantas enfermedades. La polucion es el drama y de difícil solucion
Bibliografia
Depression: the radical theory linking it to inflammation Alison Abbott considers a persuasive case for the link between body and mind
The Inflamed Mind A Radical New Approach to Depression Author: Edward Bullmore
«Después de secuenciar más de 100.000 tumores ya conocemos todos los genes implicados en cáncer»
Mi amigo el Dr Antonio Ballesta es sin duda un líder organizando congresos o similares. Tiene un alto cargo en “analizA” y he teido la suerte de asistir a alguno.
Antonio, es sencillo, andaluz y catalán y madrileño y de todas partes, buen amigo y un investigador y gestor “DE CHUPA DE DOMINE”.
El articulo sobre Mariano Barbacid, es absolutamente explicativo de los éxitos sobre el Cancer, esa maldita enfermedad que tanto hace sufrir y a la que se le está cogiendo el “tranquillo”, y convirtiéndola por lo menos en una “enfermedad crónica” y esperamos que aumente su conocimienrto y eficacia en su tratamiento
Y la razón es imprecindible siempre “buscar los orígenes”, y el próximo paso será evitar la polucion que seguramente es el principal de sus desencadenantes.
Tiene poco sentido que yo copie la publicación de Mariano Barbacid, pero me gusta hacerlo y creo que el merece que se divulguen sus conocimientos.
..Pablo Malo Segura. En las dos décadas que llevamos de siglo XXI se ha producido una auténtica revolución en las terapias que hoy día se aplican al cáncer. Hay tres grandes avances: medicina de precisión, inmunoterapia e inmunoterapia celular (CAR-T cells). Los inicios de la medicina de precisión se remontan a 1998, cuando se aprobó Herceptina, un anticuerpo monoclonal dirigido contra el receptor HER2. «Desde entonces se ha avanzado muchísimo, sobre todo, gracias a la ultrasecuenciación. Hoy en día podemos decir que después de secuenciar más de 100.000 tumores ya conocemos todos los genes implicados en cáncer, tanto oncogenes como genes supresores». Así lo ha señalado el Dr. Mariano Barbacid, profesor AXA-CNIO de oncología molecular del Centro Nacional de Investigaciones Oncológicas (CNIO), durante las IV Jornadas Científicas de Biomedicina, organizadas por Analiza.
Para el bioquímico este es un gran avance, pero a su vez es un reto importante. «Por el momento casi todos los fármacos de los que disponemos son inhibidores de quinasa, inhibidores enzimáticos, y hay un gran porcentaje de genes mutados en cáncer que todavía la ciencia no sabe cómo inhibirlos». Según ha explicado, la medicina de precisión consiste en el desarrollo de fármacos selectivos contra aquellas proteínas producidas por los genes mutados en cada tipo tumoral, independientemente del órgano de origen o las características del tumor.
«Uno de los grandes retos de la medicina personalizada es que conocer el gen no es suficiente para desarrollar un fármaco», ha precisado. Además, ha identificado otros dos grandes retos. «El primero, que la gran mayoría de los genes de cáncer no sabemos cómo inhibirlos sino son enzimas. El segundo, son las resistencias que aparecen antes de un año». Por otro lado, el Dr. Barbacid resaltó la importancia de agilizar los ensayos clínicos y recordó que «el cáncer lo producen nuestras propias proteínas».
Medicina de precisión y estratificación molecular de los tumores El Dr. Barbacid ha destacado que para que la medicina de precisión sea aplicable y efectiva, además de la clasificación anatomopatológica de los tumores es necesario un diagnóstico molecular que permita identificar los genes mutados y realizar una estratificación de los tumores de acuerdo a este diagnóstico. «Hoy en día es absolutamente esencial que los hospitales implementen análisis moleculares que identifiquen la presencia de genes mutados, siempre y cuando para estos genes ya existan fármacos efectivos». En este sentido, ha indicado que el que te detecten un oncogen de la familia NTRK puede salvarte la vida. «El fármaco que te van a dar va a ser muchísimo más eficaz que si te dan una quimioterapia o algún otro fármaco».
«
El cáncer de pulmón es donde la medicina personalizada ha tenido un impacto mayor». En concreto, donde más se ha avanzado es el adenocarcinoma. En más de la mitad de ellos se conoce el gen iniciador y para la mayoría ya existen inhibidores. «Un inhibidor solo se convierte en fármaco cuando lo aprueba la FDA», ha recordado. KRAS es uno de los oncogenes más frecuentes dentro de los adenocarcinosmas y es uno de los más malignos. El primer inhibidor de KRAS, sotorasib, se ha aprobado en 2021, pero solo es efectivo en la mutación KRASG12C (frecuente en fumadores).
Los tumores agnósticos son aquellos cuyo tratamiento sigue determinado por la presencia de un gen mutado en lugar de por sus características anatomopatológicas o clínicas. «En las últimas dos décadas con la llegada de la ultrasecuenciación masiva se han identificado nuevos oncogenes NTRK, muy frecuentes en tumores raros ya sean pediátricos o de adultos», ha manifestado.
Implementación de la medicina de precisión en la medicina molecular
El Dr. Santiago Ramón y Cajal, jefe del servicio de anatomía patológica del Hospital Universitario Vall d’Hebron y catedrático de anatomía patológica de la Universidad Autónoma de Barcelona, ha destacado que tras la implementación de la medicina de precisión ha habido un «espectacular cambio en el pronóstico y mortalidad de los pacientes con cáncer».
El Dr. Ramón y Cajal ha asegurado que la medicina de precisión debe basarse en tres pilares: un diagnóstico anatomopatológico correcto, un estadiaje del tumor y la búsqueda de biomarcadores asociados al mismo con el fin de realizar una terapia dirigida. «En metástasis estamos avanzando lentamente y hacen falta cambios de paradigma». Según ha señalado, los motivos de que el pronóstico de la metástasis sea tan malo son la heterogeneidad intertumoral e intratumoral.
En cuanto al diagnóstico anatomopatológico es indispensable un control de calidad para disminuir el grado de discordancias, que es elevado (20%). Algunas de las razones son la falta de conocimiento y especialización de los patólogos y el déficit de estudios moleculares. «Para mejorar el día a día en diagnóstico molecular es muy importante intentar concentrar el análisis patológico clásico en centros de referencia». En definitiva, el Dr. Ramón y Cajal ha abogado por trabajar en redes y nodos moleculares. «En primer lugar, habría que digitalizar las imágenes patológicas e incrementar la telepatología y los pátologos de referencia. En segundo, implementar una tecnología NGS que permita diagnosticar múltiples alteraciones moleculares, dianas terapéuticas y factores de resistencia».
Existen 17 tipos tumorales diferentes que pueden presentar una mutación en el gen NTRK. Actualmente, hay una serie de fármacos aprobados que pueden ser utilizados para atacarlos. Asimismo, el Dr. Díaz Rubio ha recalcado la importancia de disponer de «biomarcadores que sean de tipo predictivo» para conocer si el tumor puede responder a una diana terapéutica. En este sentido, ha subrayado la importancia de utilizar la tecnología para que todos los pacientes tengan las mismas oportunidades de acceso. «En oncología se financia el fármaco, pero no se financia el biomarcador», ha lamentado. De hecho, para tener biomarcadores tienen que conseguir convenios con la industria farmacéutica al no haber una financiación específica.
La inmunoterapia es un hito que abre una nueva etapa en el tratamiento contra el cáncer. «Tenemos un sistema inmunológico suficientemente potente como para destruir los tumores. No necesitaríamos fármacos si ese sistema inmunológico estuviera activado y reconociera claramente a las células tumorales», ha enfatizado el catedrático. Según ha precisado, la inmunoterapia tiene un beneficio consistente en muchos tumores, pero no responden a ella todos los pacientes. «Tenemos que trabajar con biomarcadores que nos digan cuales son los pacientes que se van a beneficiar».
Finalmente, ha enumerado algunos de los grandes retos que existen en inmunoterapia: estudiar la expresión de PDL-1, la carga mutacional, si hay inestabilidad de microsatélite, si hay alteración en los genes repadores del DNA, firmas genómicas, la infiltración por células TILs, utilizar dos marcadores en lugar de uno, la resistencia, y la posibilidad de hacer marcadores en biopsia líquida. «La medicina multidisciplinaria es el avance más importante que hemos hecho en oncología», ha destacado.
Esto alienta al medico y sobre todo al enfermo y anima a seguir en la línea de que a la pratica de la medicina se una imprescindiblemente la investigación y además buscar el origen.
¿Por qué tantas enfermedades degenerativas incluyendo al cancer.
Porque han crecido tanto estas enfermedades desde la revolución industrial.
Tanta miasma, que nos bombardean tienen que ver con los hechos, pero como combatirlas.
Limitar la polucion es factible, pero dudo de la eficacia necesaria. Hace falta enseñar al organismo utilizar su gran capacidad * Nuestro sistema inmunológico es suficientemente potente como para destruir los tumores*
Que Dios reparta suerte, dicen los toreros cuando salen a la plaza
El mundo vive un momento en que se enfrenta a importantes desafíos como el cambio climático, sin embargo, el neurocientífico portugués Antonio Damasio considera que no hay que infravalorar la capacidad humana para resolver los problemas.
*Eso Dr Damásio hay que confirmarlo, hasta ahora estamos en manos de nuestras emociones que fabrican los sentimientos y sobre ellos solo podemos actuar con farmacología o cirugía. Si sobrevivimos a la cantidad de fatalidades que estamos sufriendo y las que están por venir, es posible que esto sea cierto. Pero hasta ahora solo es solo una suposicion, nos queda la espera.
*Y como dice Harari, pasemos del Homo Sapiens Sapiens al hombre algoritmo
Damasio (Lisboa, 1944) acaba de publicar “Sentir y saber. El camino de la consciencia” (Destino) porque, según ha indicado en una rueda de prensa virtual, tenía algo nuevo que decir, en este caso, sobre la consciencia, cuyo origen relaciona con el sentir. En esta ocasión, uno de los neurocientíficos más reputados, ha escrito practicando el arte del haiku, quedándose sólo con lo esencial, para que fuera “más inteligible y llegara a un grupo más amplio de lectores”.
*Hasta los animales unicelulares necesitan sentir para reaccionar, la cuestión es reaccionar adecuadamente a lo que conocemos y hemos aprendido y a lo que va a venir y tener una conciencia adecuada para hacerlo bien.
El director del Instituto del cerebro y la creatividad en la Universidad del Sur de California (EE.UU) habla del origen y la evolución de la mente consciente, desde las primeras bacterias, hace 4.000 millones de años, hasta la actualidad, y también del sentir y de la inteligencia.
*En mi opinión lo más inteligente que he dicho Damasio es diferenciar la emoción y el sentimiento, y esto lo había ya confirmado la cirugía cuando las lesiones frontales permiten al paciente olvidarse de su dolor al mismo tiempo que cambia su personalidad.
¿Qué es ser inteligente?
Damasio señala que existe una implícita capacidad de resolver un problema de un modo ventajoso para el propio organismo, la cual se da también en las bacterias o las plantas, que “gestionan su vida pero no saben que lo están haciendo”.
En el caso de los humanos, que somos seres conscientes, que vemos el mundo que nos rodea, ser inteligente es “hacer cosas que puedan ser beneficiosas tanto para ti, como para los otros, o para la humanidad en general”.
A la vista de los problemas que encara el mundo, podría pensarse que nos estamos volviendo menos inteligentes, pero él no está de acuerdo.
“Estamos pasando por muchas dificultades, la crisis climática es un buen ejemplo. Cuando uno abre el periódico por la mañana, en lugar de besos y abrazos, leemos sobre conflictos, sobre choques”, pero cree que “no deberíamos infravalorar la capacidad humana para resolver los problemas”.
Durante la historia ha habido períodos con “muchísimos problemas, pero ahora la gente está siendo más honesta y lo está contando” y, aunque unos días al levantarse piensa: “qué horror de mundo”, otros piensan: “en el pasado resolvimos otras cosas, por qué no ahora (…) espero que resolvamos todos esos problemas”.
*Yo no sé si nos estamos volviendo más torpes pero está claro que la actitud de un número importante de ciudadanos es inadmisible y se parece no ya a los animales sino a los demonios.
Damasio dedica en su libro, de capítulos muy breves, uno a la inteligencia artificial y se pregunta cómo funcionarían los robots si tuvieran un cierto grado de vulnerabilidad, si en eso se parecieran, aunque fuera sólo un poco, a nosotros.
El neurocientífico hace hincapié en que no está diciendo que le gustaría que las máquinas sintieran como los humanos, ellas no tienen sentimientos y “es lo último que quisiera”, además, como no tienen vida “nunca van a ser como nosotros”.
El libro trata, sobre todo, de la realidad del ser humano y de otros seres vivos que tienen sentimientos, los cuales “dieron origen a la consciencia y la obsequiaron generosamente al resto de la mente”, escribe Damasio.
Estos pueden ser primarios (hambre y sed, dolor y placer) o provocados por alguna emoción, como el miedo, la ira o la alegría, y su principal función es transmitir información, que proporcionará un comportamiento. A través del sentimiento, uno acaba siendo capaz de saber, explica a los periodistas.
A lo largo de la evolución, llegó un momento en que el sistema nervioso fue capaz de comunicarse, de interactuar con el cuerpo, y es en esa interacción cuando se tiene la posibilidad de sentir.
Hay personas que piensan que la consciencia es algo “muy misterioso” y no se puede explicar, pero es porque se basan sólo en el cerebro para tratar de hacerlo.
*Y creo que lo es , la conciencia es un misterio por lo menos a la altura de nuestros conocimientos y luchamos por entenderlo. Necesitamos unir varias dimensiones, lo orgánico y funcional con lo etéreo como es la mente
El cuerpo no puede tener consciencia sin sistema nervioso, pero este tampoco puede tenerla sin sentimientos, sin el cuerpo.
*Es posible que la conciencia sea un estado evolutivo de la complicidad de un cerebro en evolución. Hasta cierto grado de desarrollo tenemos una conciencia de supervivencia. Cuando el cerebro aumenta de tamaño y se complica ya puede tomar decisiones. Pero esto es una forma de conocer nuestra realidad. Necesitamos insertar sobre esta anatomía compleja un sentimiento de yo soy yo y puedo controlar mi medio y la única aplicación que entendernos es el alma, a la que los científicos llaman mente y hasta ahora, no entendemos su composición y como se aplica a la estructura cerebral
La consciencia “es una habilidad maravillosa que nos permite saber quiénes somos, y no todas las criaturas vivas pueden decir lo mismo”, ha permitido que haya conocimiento y, con ella, hemos podido desarrollar una inteligencia basada en el razonamiento y la creatividad.
*El científico debe admitir que ciertas condiciones no las entiende todavía es posible que los algoritmos del próximo cerebro nos la aclaren.












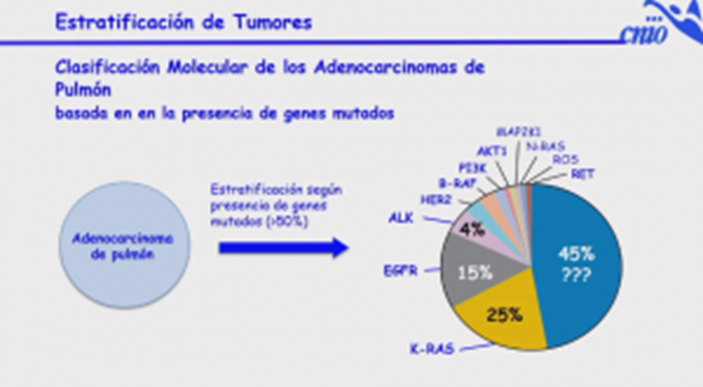
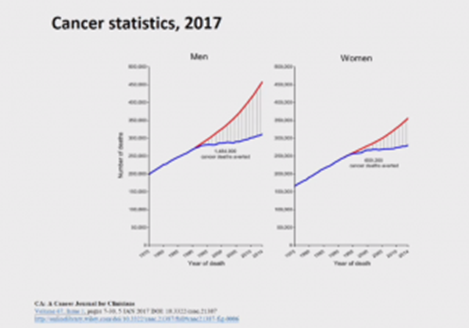

{kind=link}