
Don Santiago descubrió una hendidura en el retículo que formaba el sistema nervioso. Con ello rompió la teoría sincitial del sistema nervioso y creo la teoría de la neurona.
Tambien el sistema nervioso esta compuestos de células. “Llamadas “Neuronas” A la hendidura que descubrió Don Santiago , Sherrington las llamo Sinapsis ,
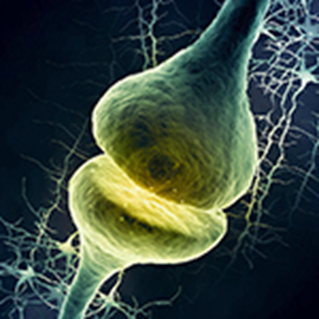
Una sinapsis es una estructura que permite a una neurona pasar una señal eléctrica o química a otra célula y asi transmitir los impulsos nerviosos de una neurona a otra y constituye la base del funcionamiento del sistema nervioso.
En una sinapsis, la neurona presináptica está muy próxima a la membrana de otra celula , la postsináptica y las separada un espacio conocido como hendidura sináptica.
La complejidad de este espacio sostienen una compleja maquinaria molecular que unen las dos membranas y llevan a cabo el proceso de señalización. En la porción presináptica destacan las proteínas implicadas en el almacenamiento, liberación y recaptación del neurotransmisor y en la postsináptica, los receptores, que son proteínas específicas a las que se une el transmisor.
En muchas sinapsis, la parte presináptica se encuentra en un axón y la parte postsináptica en una dendrita o soma. Muchas dendritas tienen expansiones de su superficie, con aspecto de chupa-chups que son característicos lugares postsinápticos y se conocen como espinas dendríticas.
Las espinas dendríticas se pueden contar con un microscopío óptico, y tener una estimación realista de los cambios en el número de sinapsis. Dado que aproximadamente el 90 por ciento de las sinapsis excitatorias se localizan en las espinas dendríticas, los cambios en su número y morfología podrían afectar a la regulación de la transmisión sináptica y a la plasticidad estructural neuronal en general y pueden estar relacionados con la comunicación cerebral atípica que muestran las personas con trastornos del espectro del autismo (TEA).
Durante el desarrollo temprano, el cerebro produce un exceso de sinapsis y posteriormente elimina aquellas que no son necesarias, no están bien o están inactivas en un proceso que se conoce como «pruning» o poda sináptica. Esas sinapsis son eliminadas en un proceso de autofagia, en el que unos elementos cerebrales comen las sinapsis alteradas o inactivas. El desarrollo postnatal de la sinapsis en la corteza cerebral de los mamíferos es un proceso dinámico que implica que la formación y la poda son simultáneas. A edades tempranas, la formación de sinapsis supera a la poda, lo que da lugar a un exceso de sinapsis excitatorias, esenciales para la estructuración de los circuitos neuronales. Posteriormente, la eliminación de sinapsis supera a la formación, dando lugar a una poda neta de espinas desde la infancia hasta la adolescencia. La densidad de espinas dendríticas alcanza su punto máximo en la primera infancia y va seguida de un pronunciado descenso durante la última etapa de la infancia y la adolescencia hasta alcanzar los niveles de los adultos. Este proceso causa la selección y la maduración de las sinapsis y los circuitos neuronales.
Los procesos de diferenciación de las células neurales y la formación de circuitos mediante contactos sinápticos entre neuronas (sinaptogénesis) ocurren en el sistema nervioso central durante las últimas fases del desarrollo prenatal y los primeros meses después del nacimiento [1,2]. Ambos procesos requieren la participación de múltiples mecanismos moleculares y celulares organizados en patrones espaciotemporales específicos, cuya alteración es la base para la aparición de anomalías funcionales, con el resultado de enfermedades psiquiátricas asociadas al neurodesarrollo. La consecuencia de alteraciones en estos procesos es una anomalía en la función de los circuitos neuronales, es decir, en el patrón de conexiones de las neuronas entre algunas regiones del cerebro o de la funcionalidad de las sinapsis entre las neuronas que conforman estos circuitos. Estas alteraciones tendrán como consecuencia un desequilibrio entre la actividad excitatoria (incremento de actividad) e inhibitoria (disminución de actividad) de las sinapsis en los circuitos afectados.

La poda sináptica en la infancia y la adolescencia Pruning
Desde que el niño nace y hasta la adolescencia su cerebro está en continua evolución. En esa etapa se generan muchas sinapsis neuronales como resultado de los aprendizajes que va adquiriendo, a la vez que se entrena la plasticidad neuronal, que no es más que la capacidad del cerebro para reorganizarse y formar nuevas conexiones neurales con el objetivo de adaptarse al entorno.
En estos años también tiene lugar un proceso muy importante en el cerebro infantil que permite reorganizar la estructura sináptica y optimizar el procesamiento de la información: la poda sináptica o poda regulatoria.
Durante los primeros años de vida, en el cerebro infantil se crean nuevas sinapsis a una velocidad sorprendente que puede alcanzar hasta las 40.000 conexiones neurales por segundo. Esto le permite al niño conocer su entorno y adquirir nuevos conocimientos a una gran velocidad, el problema es que con el paso del tiempo muchos de esos aprendizajes dejan de ser útiles. De esta manera, en el cerebro infantil se mantienen muchas sinapsis que no son funcionalmente necesarias y que en lugar de facilitar el procesamiento de la información, lo ralentizan.
Para evitar que esto ocurra y garantizar un procesamiento eficiente de la información tiene lugar la poda sináptica, que no es más que el proceso mediante el cual se eliminan las conexiones más débiles entre las neuronas que se crearon durante los primeros años de vida. Básicamente, se trata de un proceso regulador que garantiza una organización sináptica más eficiente, eliminando las estructuras sinápticas innecesarias del cerebro para aumentar la superficie de recepción de los neurotransmisores.
Por lo general, el período de poda comienza alrededor del segundo año de vida y se extiende hasta avanzada la adolescencia. La poda en las zonas de la corteza involucradas con la percepción visual y auditiva suele completarse alrededor del 4 y 6 año de vida mientras que la poda de las funciones superiores como el control inhibitorio y la autorregulación emocional continúa hasta los primeros años de la juventud.
Hasta ese momento, la poda regulatoria puede haber reducido entre un 30 o 40% el número de sinapsis en el cerebro, lo que significa que el niño habrá perdido algunas de las habilidades o conocimientos que aprendió en algún momento de la infancia y que dejó de utilizar. En su lugar se habrán creado nuevas conexiones correspondientes a las nuevas habilidades que va desarrollando.
La poda sináptica es tan importante para el desarrollo cerebral como la creación de nuevas conexiones ya que de esta manera se eliminan del cerebro las sinapsis que no se utilizan, se refuerzan las estructuras neuronales más importantes y se da paso a la creación de nuevas conexiones. Sin embargo, más allá de su importancia anatómica, la poda sináptica también beneficia el desarrollo cognitivo, emocional y conductual de los niños.
La poda al eliminar conexiones inútiles, favorece la consolidación de una red cerebral mucho más eficiente que permite que se creen nuevas sinapsis con mayor facilidad, y Contribuye a la adaptación del entorno. Durante la poda cerebral desaparecen las sinapsis que el niño ya no utiliza para dejar paso a la creación de nuevas conexiones que le permitan adaptarse con más facilidad a su nuevo entorno, como reveló un estudio realizado en la Escuela de Medicina Harvard.
Los expertos consideran la poda sináptica como un proceso madurativo en el que se consolidan las conexiones cerebrales y las funciones cognitivas más importantes en los niños. En especial, la poda de la sustancia gris, sobre todo en las áreas de la corteza cerebral, se ha relacionado con un mayor desarrollo de funciones como la memoria, el control atencional y la concentración.
La poda sináptica se ha relacionado con la creciente y rápida adquisición de habilidades y conocimientos que tiene lugar hasta avanzada la adolescencia. En cambio, las alteraciones en este proceso se han vinculado con algunos trastornos del desarrollo como el autismo, según reveló una investigación realizada en la Universidad Icesi.
La poda sináptica está fuertemente influenciada por factores ambientales y del desarrollo infantil. Básicamente, aquellas funciones que el niño necesita para desenvolverse en su entorno serán las que se conserven mientras que las habilidades que ya no le sean útiles o haya dejado de ejercitar serán las que desaparezcan. Por eso, es importante poner en práctica algunas estrategias para estimular el proceso de poda sináptica y reforzar las habilidades que se quieren preservar.
Un grupo de investigadores italianos (Pagani et al., 2021) ha publicado un artículo en la revista Nature Communications en el que señalan que hay un exceso de sinapsis en la corteza cerebral de las personas con autismo.
Muchos de los genes implicados en el autismo intervienen en el control de la traducción y la síntesis de proteínas, pero también en la estructura, funcionamiento y plasticidad de las sinapsis. Múltiples genes de susceptibilidad al TEA convergen en vías celulares que están implicadas en el sitio postsináptico de las sinapsis glutamatérgicas.
Un estudio previo (Hutsler y Zhang, 2010) había encontrado una mayor densidad de espinas dendríticas en las células piramidades de la corteza cerebral de las personas con TEA en comparación con casos control de las mismas edades. Los dos investigadores estudiaron las células piramidales de las capas corticales superficiales y profundas de los lóbulos frontal, temporal y parietal. Las mayores densidades de espinas aparecían predominantemente en la capa II de las distintas regiones corticales y en la capa V del lóbulo temporal. El aumento de la densidad de espinas iba asociado a pesos cerebrales reducidos y se encontraron con mayor frecuencia en personas con TEA con niveles más bajos de funcionamiento cognitivo
La poda sináptica es tan importante para el desarrollo cerebral como la creación de nuevas conexiones ya que de esta manera se eliminan del cerebro las sinapsis que no se utilizan, se refuerzan las estructuras neuronales más importantes y se da paso a la creación de nuevas conexiones. Sin embargo, más allá de su importancia anatómica, la poda sináptica también beneficia el desarrollo cognitivo, emocional y conductual de los niños.
También se han visto cambios en la estructura sináptica y número de sinapsis en múltiples modelos de ratón para el TEA, incluidos los modelos de ratón del síndrome de Rett, del X frágil y de la duplicidad 15q11-13, así como los ratones deficientes en los genes CYFIP1 (proteína citoplasmática que interactúa con FMR1), CNTNAP2 (proteína similar a la contactina), DLGAP2 (proteína asociada a DLG 2) y SHANK1 (dominios SH3 y múltiples repeticiones de anquirina 1). Sin embargo, no siempre son resultados en el mismo sentido, que indiquen un aumento de espinas y de sinapsis en los ratones «con autismo» frente a los animales control. La cepa de ratones C58/J muestra una baja sociabilidad, deterioro de la comunicación y comportamiento estereotipado; por lo tanto, se considera entre los modelos animales adecuados para el estudio del autismo idiopático. Un estudio de investigadores mexicanos ha analizado las diferencias en el número y morfología de espinas dendríticas en el hipocampo y la corteza prefrontal de los ratones C58/J. Los investigadores encontraron cambios en el número de espinas y en la morfología de forma dependiente de la región cerebral: una sutil disminución de la densidad de espinas en el córtex prefrontal, una mayor frecuencia de espinas de fenotipo inmaduro caracterizadas por una longitud similar a la de los filopodios o una morfología pequeña, y un menor número de espinas de fenotipo maduro en el hipocampo. Estas diferencias entre lo encontrado en estos ratones y lo observado en tejido cerebral humano ejemplifican las dificultades de utilizar modelos murinos para un trastorno como los TEA.
Las observaciones llevadas a cabo hasta el momento en personas con TEA han llevado a la hipótesis de que una poda sináptica disfuncional y una homeostasis alterada podrían contribuir a la patología del TEA. Los exámenes histológicos postmortem apoyan esta idea, ya que se ha observado repetidamente la presencia de una mayor densidad de espinas dendríticas en el tejido cerebral de los pacientes con TEA.
Las mayores densidades de espinas en las personas con TEA proporcionan apoyo estructural a la hipótesis que plantea la existencia de cambios en los circuitos sinápticos de la corteza cerebral y que estas modificaciones pueden generar un funcionamiento alterado de la actividad cerebral.
ANATOMÍA CEREBRAL EN EL AUTISMO
Genéricamente lo conocemos todos , El crecimiento del manto cerebral , sus conexiones y sus núcleos de agrupación celular, son imprescindible, para la función cerebral, que aumenta y disminuyen con la evolución.
Sin embargo lo que dificulta el entendimiento, es como estas lesiones anatómicas o funcionales proporcionan estos daños.
Es el esquema de siempre, como esta materia produce este espiritu
En el desarrollo típico, del cerebro, la corteza cerebral, se engrosa hasta aproximadamente los 2 años y luego se vuelve gradualmente más delgada hasta la adolescencia a medida que el cerebro madura. El nuevo estudio, uno de los más grandes para investigar el grosor cortical en el autismo, se alinea con otros que indican que esta trayectoria difiere en las personas con la afección.
Los cambios mas objetivos, no dicen absolutamente nada, sobre todo si los comparamos con grades cambios y lesiones del cerebro en superdotados. Hay algo mas. Pero eso lo sabíamos de siempre
Los resultados sugieren que la estructura cerebral no cambia de manera uniforme en el autismo, sino que varía con factores como la edad, el sexo y el coeficiente intelectual, Canadá. Un estudio reciente encontró que varias regiones de la corteza cerebral son más gruesas en niños y adultos jóvenes con autismo que en sus pares de desarrollo típico.
Las diferencias son mayores en las niñas, en niños de 8 a 10 años y en aquellos con un bajo coeficiente intelectual (IQ)
Conocer los procesos neurales ligados a la formación de sinapsis y circuitos cerebrales para entender su papel en las enfermedades del neurodesarrollo, como el trastorno del espectro autista (TEA) y el trastorno por déficit de atención/hiperactividad (TDAH).
Desarrollo. La actividad de los circuitos neuronales es la base neurobiológica de la conducta y la actividad mental (emociones, memoria y pensamientos). Los procesos de diferenciación de las células neurales y la formación de circuitos por contactos sinápticos entre neuronas (sinaptogénesis) ocurren en el sistema nervioso central durante las últimas fases del desarrollo prenatal y los primeros meses después del nacimiento. Los TEA y el TDAH comparten rasgos biológicos, relacionados con alteraciones en los circuitos cerebrales y la función sináptica, que permiten tratarlos científicamente de forma conjunta. Desde el aspecto neurobiológico, el TEA y el TDAH son manifestaciones de anomalías en la formación de circuitos y contactos sinápticos en regiones cerebrales implicadas en la conducta social, especialmente en la corteza cerebral prefrontal. Estas anomalías son causadas por mutaciones en genes involucrados en la formación de sinapsis y plasticidad sináptica, la regulación de la morfología de las espinas dendríticas, la organización del citoesqueleto y el control del equilibrio excitador e inhibidor en la sinapsis.
Conclusiones. El TEA y el TDAH son alteraciones funcionales de la corteza cerebral, que presenta anomalías estructurales en la disposición de las neuronas, en el patrón de conexiones de las columnas corticales y en la estructura de las espinas dendríticas. Estas alteraciones afectan fundamentalmente a la corteza prefrontal y sus conexiones.
Por lo tanto, es importante conocer los procesos neurales ligados a la actividad de los circuitos cerebrales para entender las consecuencias de su disfunción y, con ello, su papel en el desarrollo de los síntomas característicos de las enfermedades del neurodesarrollo, como son los trastornos del espectro autista (TEA) y el trastorno por déficit de atención/hiperactividad (TDAH) [3].
El TEA es una condición heterogénea caracterizada por la presencia de alteraciones del comporamiento en la interacción social y comunicación, acompañada de comportamiento estereotipado e intereses restringidos. Además de estos síntomas necesarios para el diagnóstico, el TEA a menudo se presenta con una variedad de otras manifestaciones conductuales y funcionales, como problemas de lenguaje, hiperactividad, epilepsia, déficit de atención y trastornos del sueño. El TDAH se inicia en la infancia y se caracteriza por dificultades para mantener la atención, hiperactividad con exceso de movimiento e impulsividad, y dificultades en el control de los impulsos. El TEA y el TDAH comparten rasgos neurobiológicos, fundamentalmente relacionados con alteraciones en la estructura y función de la corteza cerebral, que permiten tratarlos conjuntamente.
La actividad de los circuitos neuronales es la base neurobiológica de los procesos del sistema nervioso central que se manifiestan en la conducta y los procesos mentales (emociones, memoria y pensamiento). La función de los circuitos presenta una importante capacidad de adaptación, mediante cambios en las propiedades espaciales y temporales de las conexiones entre neuronas del circuito. Así, el cerebro construye una respuesta adecuada a los requerimientos de cada situación interna o ambiental. La base estructural de la adaptación neural es la capacidad de modificar la cantidad y la función de las sinapsis neuronales; por lo tanto, lo que definimos como plasticidad neural (neuroplasticidad) se fundamenta en la plasticidad sináptica en los circuitos neuronales [2,4]. La maleabilidad funcional se logra durante el desarrollo modulando la expresión de un conjunto de genes que regulan mecanismos moleculares y celulares que influyen en la dinámica de las conexiones sinápticas. La neuroplasticidad durante el desarrollo del cerebro presenta patrones temporales heterogéneos: existe un período crítico de mayor maleabilidad sináptica alrededor del nacimiento, que modula la regulación génica para la formación y consolidación de conexiones neuronales adecuadas mediante la influencia de los estímulos ambientales. Éstos actúan sobre un patrón de conexiones regulado por la información genética (lo que hace que los humanos generemos un cerebro humano).
TEA y el TDAH pueden ser manifestaciones de anomalías en el proceso de neuroplasticidad del desarrollo, al igual que otros trastornos neuropediátricos congénitos y adquiridos, como la encefalopatía por hipoxia neonatal, parálisis cerebral, epilepsia, distonía, discapacidad intelectual y esquizofrenia [5,6]. Desde su aspecto etiológico, ambos procesos tienen una importante carga genética, considerándose trastornos poligénicos (múltiples genes implicados con carga patogénica escasa y variable) y, por tanto, derivados de una combinación de alteraciones genéticas de novo (mutaciones espontáneas) asociadas a una predisposición derivada de variaciones comunes heredadas. Las principales anomalías genéticas asociadas a TEA y TDAH implican genes que codifican proteínas de la sinapsis [3,7].
Los conocimientos acumulados en los últimos años muestran que las enfermedades mentales de inicio en la infancia se deben a alteraciones de la formación o de la actividad de circuitos neuronales. Entre estas enfermedades destacan el TEA y el TDAH, asociados o no a discapacidad intelectual, y otros síndromes del neurodesarrollo. La aparición durante la vida temprana de las alteraciones conductuales y funcionales del TEA y el TDAH induce Es lógico pensar que las alteraciones anatómica y fisiológica están producido por una previa alteración cromosómica. Nada en nuestra biología aparece o desaparece sin la alteración genética .
En el TEA y al TDAH se modifican los procesos del desarrollo neuronal y el establecimiento de conexiones, sobrepasando la capacidad compensatoria de la neuroplasticidad del sistema nervioso central durante el desarrollo y generando alteraciones en el patrón inicial de conexiones en los circuitos neuronales.
Durante el desarrollo embrionario, los axones de las neuronas jóvenes llegan a su destino mediante procesos bien regulados de guía axonal, estableciendo conexiones inmaduras y temporales con las neuronas que están diferenciándose en las regiones diana. Durante el desarrollo de las conexiones en la corteza cerebral aparece una estructura transitoria, la subplaca, que se forma entre los 3-4 meses de desarrollo y constituye el principal compartimiento de conexión neuronal de la corteza hasta los siete meses. La subplaca desaparece progresivamente, en la etapa posnatal temprana, hasta los seis meses de vida [8]. Las fibras nerviosas que van a establecer contactos en la corteza entran primero en la subplaca y establecen circuitos sinápticos temporales, donde permanecen un ‘tiempo de espera’ antes de entrar en la placa cortical para establecer sinapsis con las neuronas de las diferentes capas corticales. Desde los siete meses de desarrollo hasta un año de vida posnatal, la subplaca es un lugar de relevo sináptico. Estas sinapsis transitorias desarrollan circuitos neuronales transitorios, que representan la base neurobiológica de la actividad eléctrica del comportamiento fetal y de los neonatos prematuros. Durante la etapa perinatal se extienden las fibras corticales desde la subplaca hacia las neuronas de la placa cortical (futura corteza cerebral), con lo que se inicia y progresa la formación de circuitos de conexión maduros entre áreas de la corteza cerebral. Se origina entonces una sobreproducción sináptica que permanece en la infancia, en la que los procesos de generación predominan sobre los de retracción sináptica, hasta llegar a la adolescencia, donde se invierte el patrón y se produce una poda selectiva de los contactos no funcionales (es decir, predomina la eliminación de sinapsis poco eficaces sobre la generación de nuevas) (Figura). El equilibrio entre producción y eliminación sináptica seguirá extendiéndose a lo largo de la vida y es lo que denominamos neuroplasticidad adaptativa y reactiva. Recientemente se ha podido demostrar que las neuronas de la subplaca se relacionan embriológica y funcionalmente con un núcleo cerebral cuya estructura y funciones son poco conocidas: el claustro [9]. El claustro está conectado recíprocamente con todas las regiones de la corteza cerebral, de forma muy significativa con la corteza prefrontal, y su función es muy relevante en el proceso de atención y el estado de conciencia [10,11]. Aunque no se han descrito diferencias significativas en la estructura del claustro en cerebros con TEA [12], sí se han encontrado en la subplaca [13]. El estudio de las posibles alteraciones de la conectividad entre el claustro y la corteza cerebral en el TEA y el TDAH parecen un prometedor proyecto para entender su fisiopatología.
En pacientes con TEA y TDAH se han descrito alteraciones del desarrollo inicial de las sinapsis en los circuitos de conexión entre áreas corticales de procesamiento complejo (que reciben y procesan de forma combinada información multimodal), sobre todo de los lóbulos frontal, temporal y parietal [7,13,14]. El proceso de sinaptogénesis está regulado por múltiples factores genéticos y epigenéticos (ambientales), por lo que corre un alto riesgo de ser alterado en el período perinatal, durante su etapa de mayor maleabilidad, dando como consecuencia trastornos del neurodesarrollo. En relación con el carácter poligénico del TEA se han descubierto genes cuyas mutaciones producen alteraciones sinápticas que cursan con TEA y TDAH, así como discapacidad intelectual y trastornos neuropsiquiátricos. Entre los genes descritos están los que codifican proteínas de organización sináptica, que incluyen complejos de adhesión celular y factores secretados [15]. Muchas proteínas codificadas por genes de riesgo para padecer TEA, TDAH o discapacidad intelectual participan en diferentes procesos de conectividad neuronal en la sinapsis, incluyendo sistemas proteicos relacionados con receptores para neurotransmisores, como el glutamatérgico (p. ej., GRIN2B), el gabérgico (p. ej., GABRA3 y GABRB3) y el glicinérgico (p. ej., GLRA2), pero también en los mecanismos de neuritogénesis (p. ej., CNTN), el establecimiento de las sinapsis (p. ej., cadherinas y protocadherinas), la conducción neural (CNTNAP2) y la permeabilidad de las membranas neuronales a iones (CACNA1, CACNA2D3 y SCN1A) [2,3]. Algunas de estas proteínas están directamente involucradas en la actividad y la formación de las sinapsis, como las neurexinas (NRXN) y las neuroliginas (NLGN). Otras proteínas forman parte de los andamios necesarios para el posicionamiento de moléculas de adhesión celular y receptores de neurotransmisores en la sinapsis, por ejemplo, los genes SHANK (SHANK1, SHANK2 y SHANK3) [3] y los que codifican las proteínas de la familia Rho-GTPasas [16]. Estas proteínas se unen en grandes plataformas moleculares de interacción con receptores de glutamato y actina asociada a proteínas, afectando de forma muy evidente el desarrollo y morfología de las dendritas. Síndromes del neurodesarrollo asociados frecuentemente con la aparición de TEA, como el síndrome X frágil, presentan anomalías importantes en la estatura de las espinas sinápticas en las dendrítas de las neuronas corticales. La distribución heterogénea en intensidad y localización de estas alteraciones en las conexiones neuronales de la corteza explicaría las diversas manifestaciones clínicas, tanto de la entidad diagnóstica (TEA, TDAH, discapacidad intelectual, etc.) como de las diferencias entre individuos con el mismo diagnóstico. EL CANTO DE LAS NEURONAS
DISPOSICION DE LAS ESPINAS DENDRITICAS
La corteza o córtex cerebral es el tejido nervioso que cubre la superficie de los hemisferios cerebrales, alcanzando su máximo desarrollo en los primates. Es aquí donde ocurre la percepción, la imaginación, el pensamiento, el juicio y la decisión. Es ante todo una delgada capa de la materia gris –normalmente tiene 6 capas de espesor–. Esta capa de células conecta con una amplia colección de vías de materia blanca, que son mucho mas numerosas y son estas preolongaciones que la conectan con todo el resto del cerebro.
La delgada capa está depositada en las circunvoluciones, lo que permite el mayor aumento numérico de estas células. Si se extendiese, ocuparía unos 2500 cm². Esta capa incluye unos 10.000 millones de neuronas, con cerca de 50 trillones de sinapsis.
La neo corteza o corteza cerebral, localiza una serie de funciones del cerebro que nos hacen ser humanos.
Esta localizada en la superficie del cerebro y tiene un espesor de entre uno y cuatro mm se llama sustancia gris y está en contacto íntimo con la sustancia blanca en alusión a sus colores aproximados.
En la corteza se depositan las células nerviosas que componen la sustancia gris y la sustancia blanca son las prolongaciones de estas células que están envueltas en mielina a la que deben su color
En la corteza del cerebro se alojan las células del sistema nervioso en cantidades de miles de millones-
Para visualizar esas células hay que teñirlas. Lo que permite verlas individualmente. Estas células de la corteza cerebral se localizan en forma de capas.
Con la tinción de las neuronas se vio que tienen un cuerpo llamado soma del que salen prolongaciones. Las aferentes se llaman dendritas y las eferentes cilindroejes. Son como árboles y de hecho la palabra dendrita proviene del griego que significa árbol.
Gracias a las tinciones podemos ver estas células y sus prolongaciones, con el método de Nissen, se tiñen los cuerpos neuronales, mientras que con el método de Golgi se tiñen las prolongaciones de la neurona. Con estas técnicas no se tiñen todas la células, y si algunas, pues de lo contrario no veríamos nada a destacar.
El teñido de las células nerviosas es selectivo. Las neuronas forman un bosque tan denso, donde todo está en contacto con todo.
Las células nerviosas se comportan como un pequeño ordenador. La información llega a través de las dendritas al cuerpo celular o soma, se elabora una respuesta que a través de una prolongación generalmente más larga, el cilindroeje, y son transmitidas a la sinapsis. Aquí se pone en contacto con las dendritas receptoras, pero no directamente, sino a través de un espacio sináptico. A medida que sale información de los cilindroejes se contacta con las células vecinas. En las terminaciones presinapticas, existen unos depósitos de neurotransmisores que el “ los descarga al espacio sinaptico y desde aquí se estimula la neurona postsinaptica, y empieza el proceso de nuevo.
Cajal con su entusiasmo y su curiosidad dijo “conocer el cerebro es conocer el camino de nuestro pensamiento y de nuestras capacidades».
El estudio del cerebro nos permite conocer su creatividad y nuestras funciones superiores .
Nosotros somos nuestro cerebro y nuestra capacidad de hacer.
El conocimiento del cerebro no sólo tiene importancia en el desarrollo filosófico y científico, sino porque nos permite conocer la patología que lo afecta con frecuencia y buscar su tratamiento.
Antes de conocer un cerebro alterado necesitamos conocer su morfología normal y su función.
La ciencia de nuestro tiempo estudian insistentemente el diseño del cerebro. Su estudio nos llevarán al conocimiento de sus circuitos normales y patológicos. Y despues a su función.
Las imágenes del universo con sus enormes magnitudes, son superponibles a las imágenes de la microscopía cerebral con sus pequeñas células que se miden por milésimas de micra.
Estamos en un mundo global nadie puede trabajar sólo. Múltiples datos publicados sobre el cerebro, tienen dificultad para entenderlos. Por ello Javier de Felipe forma un grupo heterogeneo de cientificos, que de manera individual, però conjuntandose , intentan entenderlo globalmente.
Como modelo de estudio se está utilizando las células piramidales, que se puede decir son las células principales del cerebro. Y son la principal fuente de conecciones. Necesitamos conocer como esta constituido nuestro cerebro para después expresarlo matemáticamente.
La razón de este estudio son las espinas dendriticas, evaginaciones del citoplasma de las dendritas, que son las vías aferentes de la neurona.
Las células piramidales, se transforman en modelos virtuales a través de la matemática para poder manipularlos con mayor facilidad. Lo que se llama simulación por ordenador.
Las espinas dendriticas que parecen una simple prolongación, son estructuras muy complejas que están formadas por más de 500 proteínas y lo importante es que su morfología refleja su función.
Las espinas actúan como un pequeño ordenador. Su morfología refleja su función, las que que tienen cabeza grande tienen mayor capacidad de contactar, y cuando se estrechan, la corriente elèctrica, los potenciales de accion, disminuyen.
Hay unas espinas de cabeza grande, que tienen gran capacidad de conexión, que son muy estables. La combinación de varios espinas són depósitos de memoria.
Otras espìnas, son mas delgadas y son de aprendizaje
Una neurona tiene 20.000 espinas. Para ver el detalle de la célula, se introduce un micro electrodo y se inyecta una sustancia fluorescente que permite ver las dendritas y sus prolongaciones.
La neurona piramidales son las que tienen mayor número de conexiones dentro del cerebro y son responsables de las funciones cerebrales superiores.
En la enfermedad de Alzheimer las espinas dendríticas desaparecen y al ser portadoras de la memoria, se pierde esta. El conocimiento de las espinas dendríticas su formación y su función nos permitirá reparar la pérdida de esta estructura que ocurren en las enfermedades demenciales.
Hay millones de neuronas piramidales y como hemos dicho cada una de esta tiene 20.000 espinas.
El contajé de las espinas dendríticas no se hace directamente, sino por estimación. Los investigadores han fabricado herramientas que cuentan automáticamente el número de espinas en una dendrita. Hasta ahora el contajé de cada espina costaba meses, mientras que con las herramientas de que se dispone se hace en segundos.
El tamaño de las espinas es fundamental para saber la corriente que generan, por lo que hay que calcular el volumen y construirlas en tres dimensiones. Se está intentando obtener los valores de lo volúmenes de forma automática.
La disposición de las espinas en un rosal es similar a la disposición de las espinas dendríticas, aunque las espinas del rosal son distintas pues son puntiagudas y de base ancha. Se intento disponer las espinas del rosal de varias formas espacial y buscar la fórmula matemàtica que expresara su disposición. Los intentos de ver la disposicon en un rosal, fallaron, El contaje y disposicion de las espinas, no se puede expressar matematicamente, ni de forma lineal ni elíptica, por lo que se intenta convertirlas en notas musicales y asi darle una disposicon contable.
Los parámetros que componen la disposición, orientación, longitud, forma, disposición de las espines asi como las corrientes de accion, se las convirtio en notas musicales,.
Lo que a conseguido avier de Felipe, es considerar notas musicales las distintas espinas dendríticas, de una dendrita, de una células piramidales. Se basa para ello en varios parámetros y así construye una melodía, que sin ser excesivamente precisa nos puede dar un patrón matemático de estas dendritas fundándose en su sonoridad.
Un programa matemático donde se convierten en notas musicales un fragmento de dendrita piramidal,los filopodios se han convertido en notas musicales y un arreglo musical, permite obtener una sonoridad bastante clara al interpretarla.
Pone una nota a cada espina y una vez arreglada escuchar su melodía. Ya que visualmente no es interpretable.
El sonido se repite como un morse, se repite en notas con intervalos de silencio
Esto proporciona el sonido de una sola dendrita, y hay que imaginar como suenan todas las espinas de todas las dendritas. Por lo pronto esta técnica es muy limitada pero muy significativa. Y tiene cierta belleza.
Estudia grupos de dendrita de un cerebro de 80 años y otro de 40..
En el cerebro de un paciente de 40 años tiene gran riqueza riqueza de notas, porque tiene muchas espines dendriticas. Por el contrario en el cerebro de un paciente con 80 años, las dendritas tienen menos espinas y por tanto menos sónoridad.
Con ello se ha construido una neurona virtual, que ha estudiado un ordenador neuromorfico.
Cuando se activan determinadas espinas dendríticas, los resultados sonoros son distintos a cuando es estimulan otras distintas. Esto hace que dependiendo de la abundancia de espinas, los estímulos que llegan al cuerpo celular sea diferente. Y por consiguiente los resultados seran diferentes.
Una células piramidal es como el conjunto de muchos ordenadores.
El problema es cómo utilizar todas estas dendritas, sus espinas e integrarlas en un ordenador.
Los ordenadores neuromorfico están adecuados a la faena de contar y analizar espinas , ya que los ordenadores ordinarios está muy lejos de tener esta potencia.
Nuestro cerebro al funcionar consume solamente 12 Watios y un superordenador, consume cientos de miles de watios
Aplicando este estudio a la enfermedad de alzheimer. Donde se encuentran dos tipos de proteínas anormales. La beta amiloide y la proteína tau y estudiar cómo afecta patológicamente cada una de esta proteínas.
El arte nos ayuda también entender la patología de Alzheimer. El ejemplo lo proporciona el pintor William que pintaba autorretratos y que en 1967 empezó a padecer la enfermedad de Alzheimer. Esta enfermedad empieza en el hipocampo, donde se alojaba la memoria inmediata, y desde aquí se expande a otras zonas afectando de forma próxima funciones. Este pintor fue haciendo autorretratos a medida que evolucionaba su enfermedad y es dantesca la deformacion de las imagenes que el va obteniendo de como se ve a si mismo.
La enfermedad de alzheimer es progresiva y durante unos xx años permanece con clínica suave con un deterioro cognitivo leve, hasta que el deterioro se hace marcado y muy evidenciable. De forma que morfológicamente se puede ver en los cortes de cerebro de una necròpsia en pacientes con Alzheimer, la extensión y evolución de la enfermedad y relacionarlo claramente con su deterioro cognitivo.
Desde que empieza la enfermedad histologicamente, hasta que se manifiesta en su periodo de estado clinicamente. Pasan los años suficientes, como para aplicar la pobre teràpia de que disponemos, , con la posibilidad de que en estos estadios iniciales de la enfermedad, fueran efectivas. Tenemos muchos años para introducir terapias que eviten la evolución del deterioro.
El tener marcadores biologicos para esta y su deteccion precoz, permitiria al menos lentificar la enfermedad , en tanto que no aparecen teràpias mas efectivas.
La actividad de detereioro socialment invalidante tarda mucho tiempo a partir de las primereras lesiones histologicas en los lobulos temporales.
La pregunta es porque determinada afectación cerebral no produce deterioro y si aparece cuando existe una evolución patológica de la lesiones.
El estudio se está haciendo inyectando en cerebros normales y patológicos uno marcadores que permiten reconocer como las placas de la enfermedad de alzheimer afectan a las espinas dendríticas. Se trata de ver cómo la placas amiloides afectan a su entorno.
Se ha visto claramente que la placa amiloide bloquea claramente las espinas dendríticas y empobrece de espinas las dendritas de estos implantes vitales para la memoria y el aprendizaje. Se ve claramente que alrededor de una placa las espinas desaparecen marcadamente. Las espinas se torna más delgadita y menos numerosas. Las placas mutilan las neuronas y la desconectan del resto.
El estudio de cómo actúa la proteína tau se hace con un sistema similar inyectando una sustancia fluorescente que tiñe de rojo el cerebro y permite ver si tiene o no, la tau. Ya que puede ver el deposito patologico de esta proteina.
De forma que la proteína TAU no produce disminución de las espinas dendríticas, por lo menos al principio.
A partir de aqui Javier de Felipe con la colaboracion de investigadores y sobre todo de musicos, han hecho partituras en preparaciones de pacientes con tau positivo y Tau negativo. La conversión en partitura muestra que los pacientes con tau positivo tiene más silencios y los volúmenes son más pequeños, mientras que la preparaciones sin TAU, son mas sonoras.
La música está sirviendo para para obtener datos que no se ven a simple vista
Cuando aumenta el depósito de la Tau, se pierden las espinas claramente. Y por tanto se pierden las comunicaciones entre neuronas.
Luis Buñuel que también padeció esta enfermedad, tuvo la suficiente claridad de idees, para decir: Hay que haber comenzado a perder la memoria aunque sólo sea retazos, para darse cuenta que la memoria es la que constituye nuestra vida. Nuestra memoria es nuestra coherencia, nuestra razon, nuestros sentimientos, si en ella no somos nada.
Un cuarteto intèrpretò la lectura de las música obtenida de las espinas dendríticas. Con un exito extraordinario y puede ser oido en You Tube.
La analogía entre el macro cosmo y el microcosmos, tienen tal similitud, que permite la confusión. Multiples pequeños corpusculos en el caso del cerebro, o múltiples inmensos corpusculos en el caso de los planetas que estan enlazados por fibras. Las imagenes, salvando las magnitud, permite la confusion.
Deduzco otra vez que existe un patrón de forma en el Universo para lo grande y lo muy pequeño.
La similitud enorme de distintos corpusculos, rodeados de halos, parecen los mismos modelos pero a distintas escalas.
Cabe una pregunta, si las forma es tan parecida, lo seran tamnbien las funciones. Funcionarà un cerebro como un universo
Kepler tambien musicalizo el universo de manera similar a como lo hace de Felipe con las dendritas y aqui añado el relato:
Harmonices mundi ( La armonía de los mundos , 1619 ) es un libro escrito por Johannes Kepler en la ciudad de Linz . El libro contiene la primera formulación de la tercera ley del movimiento planetario .
A Harmonices mundi Kepler intenta explicar los movimientos planetarios con base en un modelo geométrico de proporciones entre diferentes poliedros relacionando estos con escalas musicales. En esta obra muestra sus intentos de fijar las órbitas de los planetas en el interior de poliedros perfectos, o sólidos platónicos , tal como había hecho en una obra anterior, misterium Cosmographicum . Para gran decepción suya la teoría nunca funcionó y después de haber expuesto en largas páginas en esta obra la abandona finalmente mostrando que es incompatible con las observaciones y las leyes del movimiento planetario deducidas en Astronomía Nueva . Kepler intentó describir estos movimientos postulando una fuerza similar al magnetismo que él pensaba emanaba del Sol .
Kepler expuso en esta obra su teoría de que cada planeta produce un tono musical durante su movimiento de revolución alrededor del Sol y que la frecuencia del tono varía con la velocidad angular de los planetas. Algunos planetas producen notas musicales constantes: por ejemplo la Tierra sólo varía un semitono con una proporción de 16:15 (o equivalentemente la diferencia entre una nota mi y uno hace entre su afelio y su perihelio ) y Venus varía en un intervalo más reducido de 25:24. Kepler explica su razonamiento para deducir el reducido espacio de tonos propio de cada planeta en términos esotéricos.
« La Tierra canta Mi, Fa, Mi: se puede deducir de estas sílabas que en nuestro hogar podemos esperar mí seria y hace m. »
En momentos muy poco frecuentes todos los planetas podrían tocar juntos en perfecta concordancia. Kepler propuso que esto podría haber pasado una única vez en la historia, quizás en el momento de la creación.
En un libro anterior Astronomía nueva , Kepler había escrito las dos primeras leyes del movimiento planetario . La tercera ley, que indica que el cubo de la distancia media del planeta al Sol es proporcional al cuadrado de su período orbital, aparecía por primera vez en el capítulo 5 de este libro después de una larga discusión en astrología .
Bibliografía:
Bedford S.A. et al. Mol Psychiatry 25, 614-628 (2020) PubMed
El Human Brain Proyet, del que Javier de Felipe es el director está dividido en 11 subproyectos. Mas de 150 laboratorios 26 países donde más de 800 sabios intentan , conocer la complejidad del cerebro humano.
[1]Barón-Mendoza I, Maqueda-Martínez E, Martínez-Marcial M, De la Fuente-Granada M, Gómez-Chavarin M, González-Arenas A (2021) Changes in the Number and Morphology of Dendritic Spines in the Hippocampus and Prefrontal Cortex of the C58/J Mouse Model of Autism. Front Cell Neurosci 15: 726501.
Hutsler JJ, Hong H (2010) Increased dendritic spine densities on cortical projection neurons in autism spectrum disorders. Brain Res 1309: 83-94.
Pagani M, Barsotti N, Bertero A, Trakoshis S, Ulysse L, Locarno A, Miseviciute I, De Felice A, Canella C, Supekar K, Galbusera A, Menon V, Tonini R, Deco G, Lombardo MV, Pasqualetti M, Gozzi A (2021) mTOR-related synaptic pathology causes autism spectrum disorder-associated functional hyperconnectivity. Nat Commun 12(1): 6084
El blog de José Ramón Alonso. Neurociencia Exceso de sinapsis en las personas con TEA