Hace ya unos años que se está intentando hacer biopsias tumorales de manera incruenta, primero fueron las biopsias líquidas y ahora está haciendose el análisis de los olores.
Toodo lo que sea abreviar y perfeccionar, sera bienvenido.
Lo que hace falta adaptar es nuestra capacidad de asimilar y utilizar l nuevas tecnologias.
Ya no es valida la frase de : Cualquier tiempo pasado fue mejor. Simplemente fueron y ademas necesarios para llegar a nuestro tiempo.
ESTE ES EL PROTOTIPO DE SNIFFPHONE.
SniffPhone, actualmente en su fase prototipo, permite el diagnóstico precoz de cáncer gástrico a partir de la respiración exhalada de una persona. El nuevo método puede revolucionar la detección del cáncer en todo el mundo. VTT ha participado en el desarrollo de prototipos y conceptos de SniffPhone con otros nueve socios del proyecto.
SniffPhone es un pequeño dispositivo de sensores desarrollado para la detección del cáncer que se puede conectar a un teléfono inteligente. En la práctica, el usuario sostiene el dispositivo delante de su boca y exhala en el sensor para dar muestras de aliento. El dispositivo mide los compuestos orgánicos volátiles (COV) contenidos utilizando sensores químicos basados en nanotecnología altamente sensibles. Las mediciones se envían a través de Bluetooth utilizando un teléfono inteligente a una plataforma en la nube dedicada, donde son analizadas por el personal médico adecuado.
El nuevo método de detección del cáncer tiene muchas ventajas sobre los métodos tradicionales: el dispositivo es cómodo e indoloro de usar. Además, proporciona una alternativa simple, rápida y rentable para la detección de cánceres gástricos. En el proyecto, los prototipos de SniffPhone han sido desarrollados y validados mediante, por ejemplo, estudios clínicos. El siguiente paso en el proyecto es encontrar financieros para este tipo de nuevo método de detección del cáncer. Está previsto que la comercialización del dispositivo se lleve a cabo a través de una empresa derivada.
El papel de VTT en el proyecto fue la implementación de la plataforma para transferir datos desde el teléfono inteligente a un espacio de almacenamiento basado en la nube. VTT también ha estado construyendo herramientas y métodos de análisis para reconocer pacientes de alto riesgo. Además, VTT desarrolló una aplicación móvil que guía al usuario en la entrega de una muestra de aliento y proporciona al usuario un análisis preliminar de la muestra. Además, se ha desarrollado una herramienta de análisis médico para mostrar los resultados de análisis de muestras de aliento. VTT también se encargó de la implementación de la investigación e innovación responsables en el proyecto mediante métodos tales como involucrar a los usuarios finales y partes interesadas en el trabajo de desarrollo a través de entrevistas y talleres e integrar prácticas de diseño responsables en las operaciones del proyecto.
El proyecto SniffPhone forma parte del Programa de Financiación Horizonte 2020 de la UE. El consorcio del proyecto está formado por nueve socios: el coordinador Technion – Israel Institute of Technology (Israel), NanoVation (Israel), Cellix Ltd (Irlanda), microfluidic ChipShop (Alemania), University of Latvia (Letonia), University of Innsbruck (Austria), Siemens (Alemania), JLM Innovation GmbH (Alemania) y VTT.
El 21 de noviembre de 2018, el proyecto SniffPhone fue galardonado con el Premio a la Innovación 2018 por la Comisión Europea al Proyecto Más Innovador. VTT implicada en el desarrollo de un dispositivo de detección del cáncer
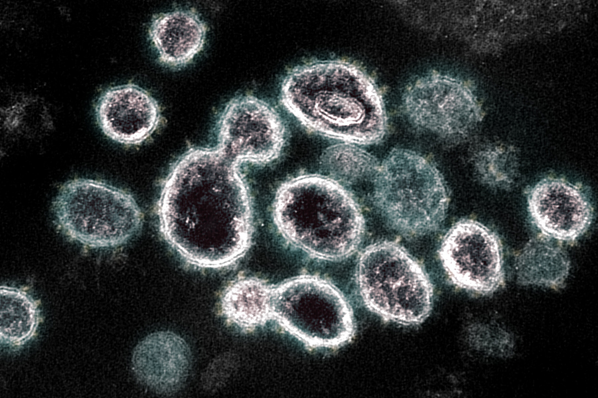
LA INTEGRINA-BETA 1 PROPAGA EL CORONAVIRUS EN EL ORGANISMO
Los virus necesitan una puerta para penetrar las células que invaden. El virus del Covi 19, posiblemente como muchos otros tienen puertas de entrada en la célula, de hecho entra por las mucosas y no por la piel, pero posiblemente tienen varias puertas de entrada.
El conocimiento y bloqueo de estas puertas de entrada daría lugar a nuevos antivirales, de los que estamos muy necesitados.
MOLECULAS DE ADHESION Y PIEL
Las moléculas de adhesión celular son glicoproteínas que se encuentran en la superficie de la mayoría de las células, median la adhesión célula a célula o la adhesión de la célula con la matriz extracelular Por ser receptores fluctúan entre estados de alta y baja afinidad con sus respectivos ligandos, los que tienen características de especificad para cada molécula de adhesión.
Todas las moléculas estructuralmente tienen un dominio extracelular, un dominio transmembrana, y un dominio intracelular El dominio extracelular en ciertas moléculas se desprende de la célula y se solubiliza en el suero, como en el caso de las selectinas.
Las moléculas de adhesión al unirse a su ligando o receptor específico, producen un cambio conformacional en el dominio extracelular que afecta la función de las células, produciendo cambios intracelulares en el citoesqueleto o en su composición química. Esto puede ocurre como una respuesta fisiológica o una respuesta patológica. Las moléculas de adhesión además están involucradas en la embriogénesis, crecimiento celular diferenciación celular etc.
Las moléculas de adhesión comprende cuatro grandes familias:
a. receptores de la familia de integrinas,
b. receptores de la superfamilia de inmunoglobulinas,
c. receptores de la familia de las selectinas y
d. receptores de la familia de las cadherinas.
Algunos autores consideran que son seis familias e incluyen a los receptores del ácido hialurónico o isoformas de CD44 y a los receptores de la proteína tirosinfosfatasa3-4.
FUNCIONES DE LAS MOLÉCULAS DE ADHESIÓN
Presentación antigénica y activación linfocitaria
El linfocito T reconoce mediante su receptor específico (TCR), al antígeno de la célula presentadora de antígenos (CPA). Esta unión es altamente específica, pero de baja afinidad y debe ser estabilizada mediante moléculas de adhesión. Una de las interacciones más importantes es mediada por LFA-1, que es una integrina y por sus ligandos ICAM-1, ICAM-2 e ICAM-3 que pertenecen a la superfamilia de la inmunoglobulinas. También interaccionan otras moléculas como CD2/LFA-3, CD28/B7 y VLA-4/ VCAM-1, para lograr una adhesión adecuada.
Las moléculas de adhesión no sólo son sistemas de anclaje, sus interacciones adhesivas modifican la conducta biológica de las células, así por ejemplo LFA-1, W-2, CD28, CD44 e ICAM-3 transducen señales intracelulares y son coestimulatorias para la producción de Interleuquina 2 (IL2), para la expresión de antígenos de activación y la proliferación de linfocitos después del reconocimiento antigénico.
Científicos Belgas, han descubierto la unión a la integrina beta-1 promueve la entrada del virus en la célula
«La unión a la integrina beta-1 promueve la entrada del virus en la célula. Una vez dentro, la célula sirve de fábrica para construir nuevos virus y luego esos virus pueden propagarse para infectar otras células u otros individuos»,
Científicos de la Universidad católica de Lovaina (Bélgica) han descubierto que la integrina-beta 1 es una de las múltiples llaves que esconden las células y que utiliza el coronavirus para introducirse y propagarse por el cuerpo humano.
Para atravesar la membrana plasmática que recubre las células, el coronavirus utiliza las proteínas presentes en las células como «cerraduras» que logran abrir para introducirse, reproducirse e infectar al resto, comenzado así el proceso de infección vírica.
Para llegar a esta conclusión, los científicos emplearon la técnica microscopía de fuerza atómica, que consiste en fijar un solo virus en el extremo de una caña de pescar y lanzarlo sobre la superficie de células vivas y posteriormente ejercer una fuerza para separar el virus de la superficie en caso de que se adhiera, comenta el experto. Así, los científicos observaron que la integrina beta-1 funcionaba de receptor e interactuaba con el virus.
El ingenio de los investigadores roza la brujeria, o por lo menos un alto poder imaginativo
Tras cinco años de investigación se abre la puerta al desarrollo de tratamientos que impidan que esta «cerradura» sea activada por el virus.
«Cada descubrimiento de una nueva llave abre nuevas posibilidades para contrarrestar el virus, por ejemplo, intentando bloquear esta nueva puerta y ver que moléculas pueden bloquearla», detalla Alsteens.
Queda, por tanto, ver las infinitas posibilidades de este descubrimiento que podría incluso ayudar a lograr tratamientos más precisos contra el cáncer, en concreto, las conocidas como terapias con virus oncolíticos.
Estos tratamientos consisten en la introducción de un virus genéticamente modificado directamente en el tumor del paciente, infectando las células cancerosas y dejando ilesas las sanas. En estos casos, la integrina-beta 1 serviría para facilitar la introducción de estos combatientes.
Bibliografía.
David Alsteens, profesor del Instituto de Ciencia y Tecnología Biomolecular de la Universidad de Lovaina .
Moleculas de adhesion y piel sanguineti ac1, Rodríguez-tafur jm2.
Entender el cambio de cadenas de ADN a ARN, es de tal dificultad, que cuesta un esfuerzo enorme comprender como los investigadores llegan a estas maravillosas conclusiones
La replicación del DNA es parcial con cada cadena parental actuando como molde para la síntesis de una nueva cadena complementaria.
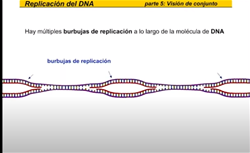
En primer lugar el DNA se desenrolla y se rompen los puentes de hidrógeno entre las dos cadenas este proceso es ayudado por la enzimas a las proteínas en las antes a cadena simple SSB y evitan que las cadenas se vuelvan a unir, esto crea una burbuja de replicación las. LAS burbujas de replicación se forman en múltiples lugares a lo largo de la molécula de DNA aumentando considerablemente la velocidad de la replicación.
mIRAB más de cerca a un extremo de la burbuja de replicación
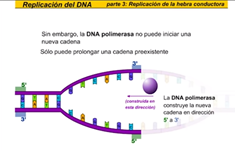
Esto es una horquilla de replicación una vez que las cadenas han sido desarrolladas y separadas la DNA polimerasa puede comenzar a construir
una nueva cadena la hebra conductora es la nueva cadena que crece de modo continuo hacia la horquilla de replicación la DNA polimerasa construye la nueva cadena en dirección 5 prima 3 sin embargo la DNA polimerasa no puede iniciar una nueva cadena, solo puede prolongar una cadena preexistente las primas a colocar los primeros
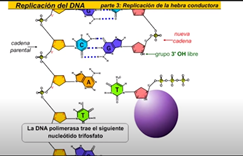
nucleótidos de la nueva cadena el segmento resultante deRNA cebador proporciona un extremo 3 prima libre al que enlazarse la DNA polimerasa puede ahora ir colocando los nucleótidos complementarios a medida que se desplazan a lo largo de la cadena molde obsérvese que la DNA polimerasa lee la cadena molde en dirección 3 prima la hélice continúa desarrollándose y abriéndose permitiendo a la hebra conductora crecer de modo continuo en la dirección de la horquilla de replicación más tarde un tipo diferente de DNA polimerasa reemplaza el cebador de rn por tn como se forma la nueva cadena de DNA veamos el proceso con más detalle echemos un vistazo más de cerca a la construcción de la nueva cadena de DNA la DNA polimerasa 3 trae el siguiente nucleótido trifosfato Enlos tres grupos fosfato la energía es liberada cuando se rompe este enlace esta energía se usa para polinizar la nueva cadena de DNA la polimerización es el proceso por el que se forman las nuevas cadenas
El proceso de acción otra vez la energía se libera el fosfato se une al grupo h libre los puentes de hidrógeno se forman entre los nucleótidos así es cómo se polimeriza las nuevas cadenas de ADN .
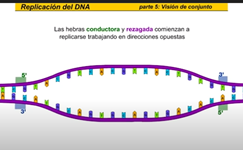
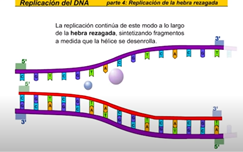
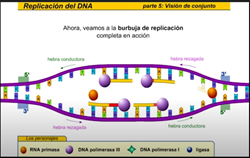
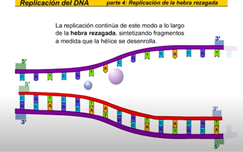
La hebra rezagada se sintetiza en dirección opuesta a la del avance de la horquilla de replicación la hebra rezagada es la nueva cadena que crece de modo discontinuo alejándose de la horquilla de replicación en primer lugar laRNA primas a añade un fragmento deRNA cebador entonces la DNA polimerasa comienza a sintetizar la nueva cadena de DNA antes de que pueda continuar la síntesis de la hebra rezagada la hélice debe continuar desarrollándose así la hebra rezagada se sintetiza de manera discontinua una vez más la RNA primas a la nueva cadena los tramos discontinuos se denominan fragmentos de Okazaki al igual que en la hebra conductora una DNA polimerasa diferente cambie RNA x DNA esta ADN a polimerasa cambia el RNA x de n y entonces una ligazas sellan la unión de los fragmentos de TN la replicación continúa de este modo a lo largo de la hebra rezagada sintetizando fragmentos a medida que la hélice se desenrolla la nueva cadena es una copia exacta de la otra cadena parental fijémonos ahora en el conjunto de la burbuja de replicación las hebras conductora y rezagada comienzan a replicarse trabajando en direcciones opuestas mientras tanto otra hebra conductora se está replicando sobre la cadena opuesta de la burbuja hay una segunda hebra rezagada en el extremo opuesto y ahora una segunda DNA polimerasa añade desoxirribonucleico cambiando los fragmentos deRNA x DNA finalmente una liga se sella la unión de los fragmentos de pene ahora veamos la burbuja de replicación completa en acción pero este proceso continúa en ambas direcciones hasta que la molécula completa de DNA ha sido replicada hay múltiples burbujas de replicación a lo largo de la molécula de DNA las burbujas continúan creciendo hasta que llegan a unirse ahora tenemos dos moléculas completas de DNA
La bibliografía esta extraída de repetidos trabajos de Internet
Pobre señora, fue la pionera en describir la estructura del ADN, pero los investigadores varones, que no podían permitir que alguien fuera, no mas listo, sino tan listo como ellos, y sobre todo si era una mujer. No la dejaron tranquila y parece que la historia no le ha dado su merito. Yo me siento dolido por estas estupideces históricas y por ello me atrevo a defenderla.
Las mujeres dedicadas a la ciencia han sido despreciadas, por la historia. Las instituciones, sus colegas masculinos y, lo que es peor, la sociedad en conjunto las han condenado a un injusto anonimato.
Rosalind Franklin constituye un claro ejemplo de ello. Poseedora de un inusitado talento para la física detectado por sus instructores a una edad temprana, a los 17 años decide ir a la universidad para estudiar Química, Física y Matemáticas. Entusiasmada por la ciencia, había escuchado a Einstein en una de sus conferencias, y decidió, tal y como éste proclamaba, poner su vida al servicio de ella.
Hace algo más de 60 años, Watson y Crick publicaron el artículo en Nature con su propuesta de estructura para el ADN. En el último párrafo y entre otros, citaban a Rosalind Franklin y le agradecían sus resultados experimentales no publicados e ideas. Años más tarde, en el libro La doble hélice, crónica muy personal del descubrimiento de la estructura del ADN, JAMES WATSON ESCRIBIÓ SOBRE ELLA QUE EL MEJOR LUGAR PARA UNA FEMINISTA ERA EL LABORATORIO DE OTRA PERSONA. Y todavía unos años más tarde, Francis Crick escribió que, en el King’s College de Londres, donde Rosalind Franklin trabajaba, había restricciones muy irritantes –no podía tomar café en la sala de profesores de la facultad porque estaba reservada para los hombres- pero solo eran trivialidades, o al menos así lo parecían entonces.
Watson y Crick se referían a Rosalind Franklin como una “FEMINISTA QUE SE QUEJABA DE TRIVIALIDADES”. Envidia cochina.
Y, sin embargo, su propuesta de estructura del ADN se basaba en imágenes de esta molécula tomadas con la técnica de difracción de Rayos X y obtenidas por Rosalind Franklin, quizá, en aquella época, la única persona del mundo capaz de conseguirlas con una calidad tan extraordinaria. Y, de nuevo, años más tarde, tanto Watson, con reticencia y muchas dudas, como Crick, creo que más sincero y elegante, reconocieron la extraordinaria calidad como científica de Rosalind Franklin y su participación esencial e irremplazable en el descubrimiento de la estructura del ADN.
Rosalind Franklin..
Rosalind Elsie Franklin nació el 25 de julio de 1920 en Londres, segunda de cinco hermanos, tres de ellos varones, en una familia judía que llevaba cuatro generaciones dedicada a la banca. Su educación más temprana, hasta los 18 años, la recibió en varios colegios de prestigio, incluyendo una estancia en Francia con un programa que incluía, además de costura y deporte, aula de debate y, sobre todo, física y química. Vuelve a casa y aprueba el examen de ingreso en el Colegio Newnham, en Cambridge, para estudiar ciencias experimentales y, en concreto, química. Su padre no acepta la decisión de Rosalind y le retira su asignación, pero una tía, hermana de su padre, corre con los gastos y le permite estudiar en el centro elegido. No pasó mucho tiempo sin que el padre aceptara la decisión de su hija y volviera a hacerse cargo de los gastos.
En 1941, se gradúa en química y física y, de inmediato, consigue una beca para iniciar su tesis doctoral pero un año después, en 1942 y en plena Guerra Mundial, pasa a la Asociación para la Utilización del Carbón y con sus investigaciones ayuda al esfuerzo de guerra. Hizo un trabajo importante en sus estudios sobre el carbón lo que le permitió, al acabar la guerra y en 1946, defender su tesis doctoral.
Fue una científica francesa refugiada en Inglaterra durante la guerra, Adrienne Weill, la orientó en su trabajo postdoctoral y, en 1947, la animó a ir a Francia, al Laboratorio Central de Servicios Químicos del Estado, en París. Era un grupo de investigación muy activo, a la última, dinámico y, sobre todo, abierto a las mujeres, a diferencia del anquilosado y masculino entorno que conocía en Inglaterra.
Allí, en París, aprendió la técnica de difracción de Rayos X en la que se convertiría en una experta a nivel mundial y aplicaría, pocos años más tarde, a la molécula del ADN. Además, amó Paris y los bistros, la comida en el campo, la cocina, las montañas, esquiar y acampar. Hizo amigos para toda la vida y, en 1951, vuelve a casa, a Inglaterra, y consigue una plaza en el King’s College de Londres. Allí, John Randall, el director del departamento, le encarga el estudio de la estructura del ADN.
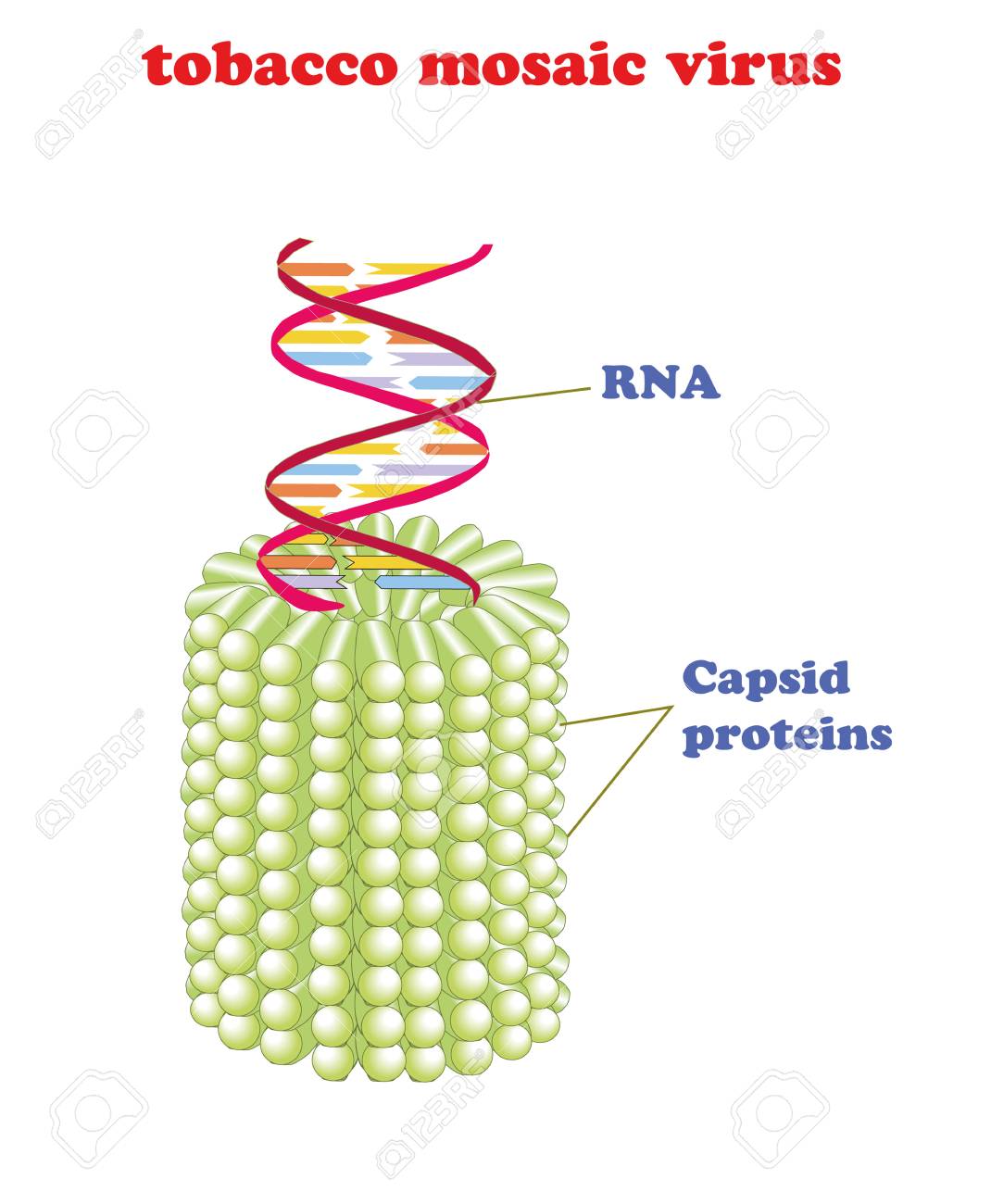
Tobacco mosaic virus with RNA isolated. Schematic vector model of tobacco mosaic virus. TMV the first virus to ever be discovered and key to gene therapy.
Pero el King’s College era, dentro del panorama machista de la ciencia inglesa, el centro que parecía encargado de guardar las esencias. Las mujeres del personal del King’s College eran tratadas con respeto pero sabían, sin ninguna duda, que nunca alcanzarían el estatus de los hombres. Era el centro en el que se mantenía, con orgullo de casta, la “trivialidad”, según Crick, de no dejar entrar a las mujeres en la sala de profesores.
En su estancia en el King’s College, Rosalind Franklin mejoró el aparato para obtener imágenes con ADN, cambió el método y obtuvo fotografías, junto a su estudiante de doctorado Raymond Gosling, con una nitidez que nadie había conseguido antes. En noviembre de 1951 dio una charla para exponer sus resultados a sus colegas del King’s College. Entre el público estaban Watson y Crick, también interesados por la estructura del ADN, y que trabajaban en el Laboratorio Cavendish, en Cambridge, a unos 90 kilómetros al norte de Londres. Era Maurice Wilkins, compañero, aunque no se llevaban bien, de Rosalind Franklin en el King’s College, y también estudioso de la estructura del ADN y buen amigo de Watson y Crick, quien les había invitado. En aquel seminario, Watson y Crick empezaron a conocer el trabajo de Rosalind Franklin y a utilizar sus datos.
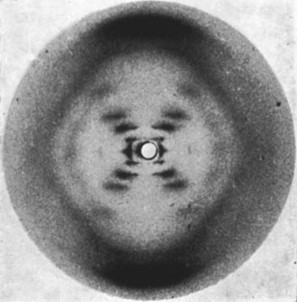
Fotografía número 51 del ADN
Fue también Wilkins quien, en los meses siguientes, fue enseñando a Watson y Crick imágenes de ADN tomadas por Rosalind Franklin, rara vez con su permiso y la mayor parte de las veces sin que ella lo supiera. En febrero de 1953, vieron tres imágenes y, entre ellas, la famosa fotografía número 51. Para entonces, Watson y Crick llevaban más de un año sin conseguir nada positivo. La número 51 la habían conseguido Franklin y Gosling en mayo de 1952 y, años después, Watson recordaba lo que sintió cuando la vio: En cuanto ví la foto quedé boquiabierto y se me aceleró el pulso. Por lo que se sabe, Rosalind Franklin nunca se enteró de que Watson había visto la fotografía.
Estas imágenes, más los datos de la charla de Rosalind Franklin de noviembre de 1951, más algunos datos más proporcionados por Wilkins, llevaron a Watson y Crick a su propuesta de la estructura del ADN y la publicaron en Nature en abril, solo un par de meses después de ver la número 51.
En el artículo, Watson y Crick mencionan a Rosalind Franklin, entre otras personas, y sin ninguna mención especial a sus datos y sus fotografías: …hemos sido estimulados por el conocimiento de la naturaleza general de resultados experimentales no publicados y las ideas de Wilkins, Franklin y sus colaboradores.. Así es de enigmático a veces el lenguaje científico, además de ser un ejemplo impagable de cómo subestimar el trabajo de otro.
En el mismo número de Nature, unas páginas más adelante, Rosalind Franklin y su doctorando Raymond Gosling, publicaron un artículo muy técnico sobre sus fotografías, con la famosa 51, y, demostrando su honradez científica, y personal, apoyando el modelo propuesto por Watson y Crick.
Hay quien ha propuesto que, para entonces, Rosalind Franklin había llegado a las mismas conclusiones que Watson y Crick, pero la rapidez de la publicación le impidió proponer su modelo. En 1951 había escrito que sus resultados sugerían una estructura helicoidal con 2, 3 o 4 cadenas y con los grupos fosfato hacia el exterior. Esto lo escribió 16 meses antes del famoso artículo de Watson y Crick.
Pero ya estaba cansada de sus discusiones con Wilkins, Watson y Crick y, en general, del ambiente del King’s College. Se traslada al Birbeck College, también en Londres, al laboratorio dirigido por John Bernal, donde permaneció hasta su muerte.
En este centro su carrera investigadora siguió adelante, con importantes trabajos sobre virus, en concreto, el del mosaico del tabaco y el de la polio que todavía citan los expertos. Pero en 1956 se siente mal durante un viaje por Estados Unidos y pronto se le diagnostica cáncer de ovario, quizá provocado por la excesiva exposición a radiaciones durante sus investigaciones con Rayos X. Todavía trabajó durante otros dos años, y después de tres operaciones quirúrgicas y quimioterapia, técnica que entonces estaba empezando a aplicarse, murió en Londres el 16 de abril de 1958, a los 37 años. Cuatro años después, en 1962, Watson, Crick y Wilkins recibían el Premio Nobel por sus estudios sobre la estructura del ADN. Ni Watson ni Crick mencionaron a Rosalind Franklin en sus discursos de aceptación.
Es curioso, pero son las tonterías que Watson hizo en los cincuenta y relató en los sesenta, las que crearon la admiración con que Rosalind Franklin es hoy recordada. Sus colegas del King’s College la consideraban demasiado “francesa”, o sea, liberal en sus costumbres, vestidos, intereses intelectuales y temperamento. Era directa y apasionada, le encantaba el debate, era seria y, a veces, dura y abrasiva. Wilkins, por ejemplo, la consideraba desagradable.
Watson, como siempre, fue el más cruel y en La doble hélice, su libro de memorias de aquella época, escribe párrafos que rozan el insulto: Estaba decidida a no destacar sus atributos femeninos. Aunque era de rasgos enérgicos, no carecía de atractivo, y habría podido resultar muy guapa si hubiera mostrado el menor interés por vestir bien. Pero no lo hacía. Nunca llevaba los labios pintados para resaltar el contraste con su cabello liso y negro, y, a sus 31 años, todos sus vestidos mostraban una imaginación propia de empollonas adolescentes inglesas. Quizá el párrafo, corto y directo, que mejor demostraba el problema de Watson en su trato con colegas científicas es aquel en que le aconseja a Wilkins que era evidente que, o Rosy se iba, o habría que ponerla en su sitio. Por cierto, nadie llamaba Rosy a Rosalind Franklin, solo Watson y Wilkins, y quizá Crick, y a sus espaldas.
El certificado de defunción de Rosalind Franklin dice: Una científica investigadora, soltera, hija de Ellis Arthur Franklin, un banquero. Nos vale como definición y como recuerdo.
Sres Watson y Crick. Os portasteis mal con Rosalin, pero que sepáis que la verdad prevalece y que todos sabemos que esta Señora , que parece que alguna vez tenia mal humor, hizo o casi hizo el mayor descubrimienro que ha tenido la historia de del ADN
Que Dios la bendiga, y a los otros, que se lo recuerde.
Algo así como “no me causan pavor vuestras palabras esquivas”
Referencias
Attar, N. 2013. Raymond Gosling: the man who crystallized genes. Genome Biology 14: 402.
Braun, G., D. Tierney & H. Schmitzer. 2011. How Rosalind Franklin discovered the helical structure of DNA: Experiment in diffraction. Physics Teacher 49: 140-143.
Crick, F. 1988. Que loco propósito. Tusquets Eds. Barcelona. 209 pp.
Elkin, L.O. 2003. Rosalind Franklin and the double helix. Physics Today March: 42-48.
Glynn, J. 2008. Rosalind Franklin: 50 years on. Notes & Records of The Royal Society 62: 253-255.
Glynn, J. 2012. Remembering my sister Rosalind Franklin. The Lancet. 379: 1094-1095.
Maddox, B. 2003. The double helix and the “wronged heroine”. Nature 421: 407-408.
Olby, R. 1991. El camino a la doble hélice. Alianza Ed. Madrid. 694 pp.
Rodrigues da Silva, M. 2010. As controversias a respeito da participaçao de Rosalind Franklin na construçao do modelo da dupla hélice. Scientae Studia 8: 69-92.
Watson, J.D. 1978. La doble hélice. Plaza & Janés Eds. Barcelona. 256 pp.
Watson, J.D. & F.H.C. Crick. 1953. A structure for deoxyribose nucleic acid. Nature 171: 737-738
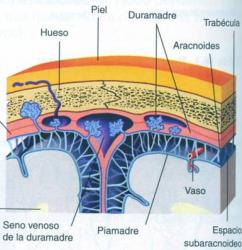
Desde la época neolítica, es frecuente la trepanación, llegando su técnica a grados insospechados de perfección en las tribus que habitaban el Alto Perú y en las del norte de Africa.
Ella se hacía de preferencia en los parietales, frontal y occipital. Se han encontrado cráneos sometidos a dos o más trepanaciones, efectuadas en condiciones satisfactorias a juzgar por el estudio de cicatrización de ellas.
En épocas más recientes, especialmente en el siglo Xix1 el conocimiento de la asepsia y de la anestesia impulsó notablemente la cirugía. Se ensayaron nuevos tratamientos cruentos contra la locura, dirigidos erróneamente a las manifestaciones orgánicas que aparecen frecuentemente en las psicopatías. La esfera sexual, las glándulas de secreción interna, fueron intervenidas quirúrgicamente con miras a suprimir Ia causa de las psicosis.
A fines del siglo pasado se localiza nuevamente la causa de las psicopatías en el cerebro y se intenta de nuevo la trepanación como tratamiento. En trabajos publicados en 1895, por Macferson, la trepanación se indicaba − a juzgar por la casuística que presenta − en enfermos que además de sus síntomas psíquicos presentaban alteraciones de Ia vista, cefaleas, vómitos, crisis convulsivas o trastornos motores. La intervención consistía en dos agujeros de trepanación, de preferencia en la zona rolándica y la abertura y drenaje de la cavidad subdural. Los síntomas regresaban ostensiblemente − a la luz de nuestros conocimientos los interpretamos como síntomas de hipertensión intracraneana por tumores probables de los lóbulos frontales que explicaban Ia sintomatologia psíquica.
Goltz y Burkhart − Director de un sanatorio suizo − experimentando en perros, pudieron apreciar que la resección de la parte anterior de los hemisferios los tornaba menos agresivos. Burkhart basado en estas experiencias sometió a un paciente que sufría de una típica esquizofrenia hebefrénica de 15 años de evolución − a repetidas intervenciones, observando sus resultados. La operación se hacía solamente en el lado izquierdo en la región temporal, parietal y frontal sucesivamente. La enferma mentalmente no se recuperó, pero quedó tranquila y ligeramente eufórica. Operó un total de seis enfermos y el gran escolio eran los trastornos motores y afásicos consecutivos a la intervención. Burkhart limitó sus aspiraciones a transformar un alienado peligroso en uno tranquilo.
EI desarrollo ulterior de Ia neurocirugía impulsado tan decididamente durante la guerra del 14 al 18, ha permitido a su vez el avance de la psico-cirugía. Las lesiones cerebrales traumáticas aceleraron el estudio de las localizaciones, ubicando con cierta precisión los polos frontales como el asiento de la conciencia
La primera vez que en la historia de la medicina, conocemos que una lesión en los lóbulos frontales, altera la personalidad del enfermo.
Sabíamos, que las lesiones cerebrales, eran incapaces de lesionar sentidos como, el lenguaje, la visión o áreas motoras en general. Pero nunca se había visto un distrés en la personalidad de un paciente.
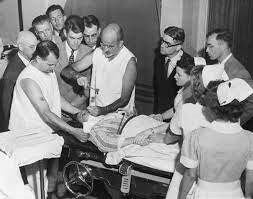
Ocurrió una tarde de verano de 1848, durante las labores de construcción de una nueva línea de ferrocarril en el estado norteamericano de Vermont. Para allanar el terreno, los barreneros introducían pólvora en el fondo de un agujero perforado en la roca, colocaban el detonante, añadían arena para taponarlo y aplastaban la mezcla con una barra de hierro. En una de estas operaciones, al capataz, Phineas Gage , se le olvida poner la arena, de tal manera que, al explotar la pólvora, la barra sale disparada y le atraviesa la cabeza de abajo arriba.
Para sorpresa de todos, el joven de 25 años no solo no muere en el acto, sino que además se recuperará de forma milagrosa, aunque sufrirá un cambio de personalidad radical. “Gage ya no fue Gage”, en palabras de su médico. Tal es su carácter que le acaban echando de todos los trabajos por falta de disciplina. Los ataques de epilepsia, secuela de la herida, se hacen cada vez más frecuentes hasta su muerte, en 1860 y además le aparece un síndrome de Falso Diogenes
La “lobotomía” accidental de Gage ha pasado a la historia de la neurología como el primer caso que documenta los efectos causados por una lesión en los lóbulos frontales del cerebro, que es la parte que se interviene en las lobotomías quirúrgicas.
La especialización cerebral
Aunque en la prehistoria ya se practicaban trepanaciones, en tiempos modernos, las primeras cirugías sobre un cerebro físicamente sano con el fin de remediar los síntomas de un trastorno o enfermedad mental –práctica conocida como psicoterapia– no se producen hasta el último cuarto del siglo XIX. Por entonces, los científicos ya habían aportado pruebas de la especialización cerebral. Paul Broca, por ejemplo, había identificado las áreas cerebrales que controlan las funciones del lenguaje, ubicadas en el lóbulo frontal izquierdo.
La observación de estos primeros trabajos llevaron al psiquiatra suizo Gottlieb Burckhardt a pensar que quizá podría paliar los desórdenes psíquicos de sus pacientes de la Maison de Santé de Préfargier aligerando su masa cerebral. La primera candidata fue una mujer de 51 años aquejada de alucinaciones, a la que, en 1889, Burckhardt extirpó una pequeña porción de corteza cerebral de unos cinco gramos por un procedimiento quirúrgico llamado topectomía.
La intervención se repitió otras cuatro veces durante los siguientes catorce meses, y, aunque las alucinaciones no cesaron, la conducta violenta de la mujer fue disminuyendo poco a poco. Entre 1889 y 1891, Burckhardt practicó la técnica con cinco enfermos más, obteniendo resultados desiguales.
Cuando, al año siguiente, presentó su informe, la comunidad médica se le echó encima, al considerar una auténtica aberración el hecho de lesionar un cerebro físicamente intacto. Tal fue el rechazo que habría que esperar casi medio siglo antes de que alguien se atreviera a repetir una psicocirugía.
Las primeras leucotomías
En julio de 1935 tiene lugar en Londres el Segundo Congreso Internacional de Neurología, en el que John F. Fulton y Carlyle Jacobsen, fisiólogos de la Universidad de Yale, presentan los resultados de unos experimentos en los que se habían extirpado los lóbulos frontales a dos chimpancés Lucy y Becky. Antes de la operación, estos animales tenían un temperamente histericos y tenían ataques de coleras cuando cometían errores al realizar tareas que se recompensaban con comida. Inmediatamente despues de la cirugía, se tornaban dociles y apacibles.
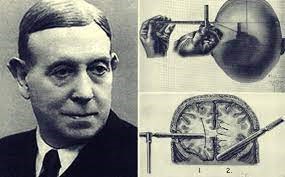
A la presentación acudieron, varios investigadores, entre ellos António Egas Moniz, de la Universidad de Lisboa, que tenia . 61 años, muy conocido por su carrera política y diplomática –había sido ministro de Asuntos Exteriores de Portugal y practico la primera angiografía cerebral como por haber descubierto la angiografía cerebral, que permitía visualizar las arterias del cerebro y por la que estuvo nominado dos veces para el Premio Nobel.
En noviembre del mismo 1935 Moniz y Pedro Almeida Lima, al que yo y en repetidas ocasiones he conocido, en las reuniones de la Sociedad Luso Española de Neurocirugia
Operan a una paciente de 63 años que presenta alucinaciones y violentos ataques de ansiedad, entre otras psicopatologías
Almeida practica dos trepanos en ambas regiones frontales del cráneo.
Inyecta alcohol puro en el interior del cerebro.
La idea era que al igual que había pasado con los monos Lucy y Becky, destruirían la zona cerebral, que elaboraba la patología de obsesiones y delirios.
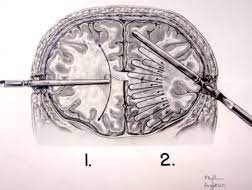
En 5 meses, intervinieron a una veintena de pacientes, introduciendo una modificación en la técnica, a la que bautizaron con el nombre de leucotomía.
Posteriormente El propio Moniz diseñó el leucotomo, un instrumento de acero en forma de estilete hueco que cortaba un trozo de lóbulo frontal, coreteza y sustancia blanca .
En 1937 publicaron un libro y trece artículos por lo que concedieron a Moniz del Nobel en 1949.
El procedimiento era eficaz en algunas personas.
Inmediatamente la comunidad cientifica argumento que no era seguro, puesto que se basaba en especulaciones y ni siquiera se habían realizado investigaciones previas en animales.
Los 20 primeros casos alcoholizados por Egaz Moniz obtuvo una recuperación franca en 7, una recuperación dudosa en otros 7 y ninguna mejoría para los 6 restantes. EI síndrome más accesible al tratamiento fué Ia agitación de los enfermos.
Nunca antes de Egaz Moniz se había intentado una intervención bilateral, aún cuando se comprendía su necesidad, en los trabajos experimentales.
La contribución de Egaz Moniz es una de las más brillantes en el campo de la psico-cirugía que abre un panorama pleno de posibilidades a la terapéutica psiquiátrica.
Siguiendo la escuela de Egaz Moniz numerosos cirujanos practican las leucotomías, modificando la técnica o sustituyendo el alcohol por novocaína, formalina.
Dr. Freeman a los 28 años, es el director de laboratorio del hospital psiquiátrico St. Elizabeths, en Washington, Walter Freeman está obsesionado con identificar alguna diferencia física entre el cerebro de los individuos psicóticos y el de los sanos, disparidades que le den alguna pista sobre la enfermedad y su cura. En 1936 lee los trabajos de Moniz y se abre un mundo ante él.
En septiembre de ese mismo año, Freeman, asistido por el cirujano James W. Watts, realiza la primera intervención para tratar un trastorno psiquiátrico en Estados Unidos. La paciente, una mujer de 63 años diagnosticada de depresión agitada, fue sometida a una leucotomía prefrontal siguiendo el procedimiento de Moniz.
Entusiasmados con los resultados favorables, Freeman y Watts repiten una y otra vez las lobotomías –término que ellos acuñan– e introducen algunas variables, como, por ejemplo, el uso de una espátula plana para realizar los cortes con anestesia local.
Pronto aparecen titulares sensacionalistas como “La cirugía del alma” o “Milagro de la cirugía”, creando una verdadera repulsión en unos y adicción a la técnica en otros.
Algunos pacientes presentaron graves secuelas la tras la lobotomía.
Freeman se entusiasmo con este empezó a practicar por su cuenta lobotomías, en masa, utilizando directamente un picahielos como instrumento.
Llegaban al hospital en un “lobotomóvil”, el personal ya tenía colocados a los enfermos en fila. Freeman insertaba el punzón metálico bajo el párpado para atravesar la cuenca de los ojos con unos ligeros golpes de martillo. Con cada martillazo, el punzón se adentraba suavemente en el lóbulo frontal del paciente, seccionándolo y destruyéndolo.
Esta técnica desplazada hoy, parece atroz se popularizó en todo el mundo .
Pero hay que situarse en la primera mitad del siglo XX, la comprensión científica de la enfermedad mental era muy limitada, por lo que no existía una cura real. El psicoanálisis no era eficaz para los grandes trastornos de la personalidad, y las llamadas “terapias somáticas”, como el electrochoque o los comas insulínicos, eran alternativas tremendamente agresivas.
Así pues, los centros psiquiátricos se convirtieron en puros almacenes de enfermos mentales, que se colapsaron, en especial, después de las dos guerras mundiales.
La leucotomía de Moniz y, más tarde, la lobotomía transorbital de Freeman eran procedimientos rápidos y baratos porque no requerían quirófanos ni material demasiado especializado. En algunos casos, además, funcionaban, por lo que se veían como una opción viable para descongestionar los centros, se practicaron 40.000 Leucotomias solo en Estados Unidos.
Entre estos trabajos se destaca el de Rizatti-Bogarello que presentó una serie de los casos y posteriormente 200 casos más. Como resultado de su experiencia insiste en el valor de la leucotomía combinada al shock cardiazólico e insulínico.
Desde 1935 a esta época la técnica de la leucotomía ha sufrido continuas variaciones, tantas que puede decirse sin exageración que cada psico-cirujano que la practica posee una técnica propia. Entre estas modificaciones merecen citarse la de Freeman y Watts de Estados Unidos de N. A. que es la que cuenta con mayores partidarios.
Aparece la clorpromazina.
Hasta que en 1954 se descubre la clorpromazina, la primera sustancia con efectos antipsicóticos, y poco a poco se abandonan las lobotomías Los fármacos revoluciónaron la psiquiatría.
A la altura de las ciencias de nuestro tiempo, la técnica permite hacer lesiones selectivas de vías fronto-talamicas con esteretoxia y mas actualmente con ultrasonidos, sin cirugía.
Los resultados, no terminan de dar resultados buenos, ya que por otra parte los tratamientos con , medicamentes si que dan unos resultados casi buenos.
Conclusiones
Estas son las conclusiones que proporciona , las cuales no creo que tengan ubicación hoy, pero por respeto a las opiniones las copio
REV CHIL NEURO-PSIQUIAT 2011; 49 (3): 225-240 Prof. Alfonso Asenjo, Dr. Isaac Horvitz, Dra. Adriana Vergara, Dr. Mario Contreras Instituto Central de Neurocirugta y Sección A de Psiquiatría del Manicomio Nacional
El análisis de las leucotomías realizadas hasta el día de hoy, nos permite sacar conclusiones de conjunto, que pasamos a resumir en los siguientes puntos:
1) La lobotomía debe practicarse en enfermos crónicos que hayan sido sometidos a los tratamientos de rigor.
2) Debe limitarse a los enfermos psiquiátricos crónicos e intentarse en las psiconeurosis sólo cuando se trata de enfermos de larga evolución que no han respondido a otras terapias.
3) El éxito de la lobotomía es más probable en aquellos enfermos que no han experimentado un deterioro mental muy acentuado y en los que no se hayan desconectado totalmente del ambiente.
4) La conducta antisocial (agresividad e impulsividad) y los síndromes angustiosos son los más beneficiados por la lobotomía.
5) Los síndromes delirantes alucinatoríos persisten en la mayoría de los casos, igual en su forma, pero disminuye enormemente su repercusión emocional.
6) La avaluación definitiva de los resultados sólo debe apreciarse por lo menos después de tres años de la intervención.
7) Los riesgos operatorios no son mayores que en cualquiera otra intervención quirúrgica (Hemorragia de la cerebral anterior).
8) Hacia el final de la intervención el paciente experimenta una desorientación total.
9) Los accidentes post-opera torios son escasos. La enuresis transitoria es casi constante. No se observan otros trastornos neurológicos de importancia.
10) En Ia evolución post-operatoria revelan frecuentemente un estado de estupor e indiferencia, cuya duración e intensidad no tienen valor pronóstico.
11) Posteriormente se observa una regresión al estado infantil (falta de control de las funciones escretoras, bulimia).
12) Gradualmente el enfermo vuelve a Ia normalidad (3-6 meses).
13)La evolución posterior depende en gran parte de Ia personalidad pre-psicótica y pre-operatoria del enfermo.
14) Debe insistirse, cada vez más, en la importancia del cuidado psiquiátrico post-operatorio (educación de los familiares, reeducación del enfermo, actividad ocupacional vigilada y graduada).
15) Los éxitos obtenidos por la mayoría de los neuro-cirujanos y los nuestros autorizan para considerar la lobotomía como tratamiento habitual de las psicosis.
BIBLIOGRAFIA
1. Allen, William F.− Results of prefrontal lobectomy on acquired and on acquiring correct conditioned differential responses with auditory, general cutaneous and optic stimuli. Am. J. Physiol. 139: 525-531. Agosto 1943.
2. Egas Moniz.−Tentatives opératoires en quelques cas de psichoses]
3. Fleming, G. W. Τ. H. y Me Kissok, Wilie.−Prefrontal leukotomy. Lancet 1: 361-362, Marzo 20-1943.
4. Freeman, Waiter.− Surgical aspects of prefrontal lobotomy, 1945.
5. Freeman, Walter.− Intelligence following prefrontal lobotomy in obsessive tension states, 1945. 6. Freeman, Walter.− Prefrontal lobotomy, 19467. Freeman, Walter y cols.− Neurosurgical treatment of certain abnormal mental states, 1944. 8. Freeman, Walter y Watts, James W.− Psico-cirugía. [ Links ]
9. Freeman, Walter y Watts, James, W.− Prefrontal lobotomv. Am. J. Psych. 101: 839-748. Mayo, 1945. [ Links ]
10. Fulton, J.− Fisiología del sistema nervioso. [ Links ]
11. Hostatter, Leopold, Smolik Edmund A. y Busch Anthony K. Prefrontal lobotomy in treatment of chronic psychoses with special reference to section of orbital areas only. Arch. Neurol, and Psvch. 53: 125-130. Febr., 1943. [ Links ]
12. Hutton E. L.− Results of prefrontal leukotomy. Lancet 1: 362־ 366. Marzo 20, 1943. [ Links ]
14. Kindwall, Josef A. y Cleveland David.− Prefrontal lobotomy. Am. J. Psych. 101: 749-755. Mayo, 1945. [ Links ]
15. Schroedder Α.− El lóbulo frontal. Montevideo 1942.
16. Lobotomias, revista Historia y Vida. Numero 608
17. REV CHIL NEURO-PSIQUIAT 2011; 49 (3): 225-240 Prof. Alfonso Asenjo, Dr. Isaac Horvitz, Dra. Adriana Vergara, Dr. Mario Contreras Instituto Central de Neurocirugta y Sección A de Psiquiatría del Manicomio Nacional.
Estás palabra fueron dichas por una paciente, al día siguiente de ser operada con una críotalamotomía para mitigar el enorme y persistente dolor que sufría como consecuencias una carcinomatosis difusa abdominal .
El dolor persistía pero ya no le importaba.
La emoción del dolor seguía, pero la interpretación , el sentimiento del mismo no era percibido.
Los sentimientos son «experiencias mentales de estados corporales«, que surgen cuando el cerebro interpreta emociones, que son estados físicos que surgen de las respuestas del cuerpo a los estímulos externos.
La emoción es un estado corporal.
Sentimiento es la conciencia que te informa de estos. Damasio
La respuesta emocional se produce cuando el individuo esta en situaciones que se oponen o favorecen sus necesidades u objetivos. El afectado se ve obligado de forma involuntaria a ejercitar funciones mentales y somáticas que no siempre tienen utilidad , al menos concreta. Las respuestas emocionales se han definido tradicionalmente como encaminadas a la proteccion del individuo o de la especie. Asi como en los animales la respuesta emocional esta estandarizada, en el ser humano son variables y matizadas por la influencia que tienen el medio cultural y la experiencia personal que ha adquirido. Describir las vías del dolor es primordial, para agregar, como la interrupción de algunas de estas vías a nivel talamico, produce, desconexión del dolor pero también a veces del sentimiento que se tienen de el.
TIPOS DE EMOCIONES
La Asociación Internacional para el Estudio del Dolor lo definió como «una experiencia sensitiva y emocional desagradable, asociada a una lesión tisular real o potencial»
Existen diferentes clasificaciones del dolor. Aquí intentamos clasificar el dolor desde un punto de vista académico según su fisiopatología subyacente (Nociceptivo y Neuropático), según su duración de presentación (agudo, crónico) y otros tipos de dolor (oncológico, psicógeno).
En la actualidad la farmacología permite controlar con bastante éxito los dolores Nociceptivo no ocurre lo mismo con los Neuropáticos.
El dolor comporta una sensación cuando se percibe y un sentimiento cuando se analiza consciente o inconscientemente.
Miguel Hernandez a la muerte de su amigo dice: Tanto dolor se agrupa en mi costado, que por doler me duele hasta el aliento.
Es la definición mas acertada que conozco .
Hace más de 100 años se empezaron a utilizar procedimientos quirúrgicos para controlar el dolor, haciendo lesiones dentro de los conocidos ciruitos del dolor.
algunos de los cuales, no quitaban el dolor y además dejaban unas enormes secuelas
Posteriormente , Egaz Moniz, empezó a tratar las alteraciones psiquiátricas, con lesión de ambos lóbulos frontales y esto dio también alguna oportunidad a la analgesia.
Mi tesis doctoral está hecha sobre lesiones de los núcleos “intralaminares del tálamo con frío”.
Si los resultados no fueron maravillosos , si mejoraban un porcentaje importante de los pacientes sobre todo del dolor nociceptivo
Las lesiones del tálamo con frío, “criocoagulación” fueron muy eficaces durante un tiempo, cuando empezó a utilizarse bomba de morfina de perfusión espinal, los ultrasonidos y la electroestimulación, las lesiones talámicas fueron desplazadas.
En la actualidad y sobre todo para el temblor , se localizan los núcleos del tálamo a tratar con resonancia magnética y los ultrasonidos, como lesivos, sin abrir el cráneo y están proporcionando resultados excelentes.
Personalmente cuando trabajaba en el hospital Virgen del rocío de Sevilla y muy jovencito tenía la cirugía ESTEREOTÁCTICA como dedicación preferente y entonces desarrollamos la cirugía talámica para el dolor lo que motivó mi afición esta técnica
Este es el fundamento de mi tesis doctoral
La analgesia que producen las lesiones de los núcleos intralaminaares del tálamo
Concretamente la lesión bilateral y simétrica de los núcleos del tálamo, Centro Mediano, (CM), Parafascicular (PF) y Dorso Medial (DM), produce una analgesia en los dolores intratables con procedimientos medicos y quirurgicos.
El fundamento de esta técnica.
Es conocido desde los años 1960 por los trabajos de Hassler, que la lesión de los núcleos somatosensoriales del Tálamo. Ventral Postero Lateral (VPL) y Ventral Postero Medial (VPM), producían dos tipos de alteraciones:
a.- Una dramática perdida de todas las sensibilidades contralaterales y la aparición en un alto porcentaje de los enfermos de disestesias contralaterales insufribles para el enfermo.
b.- Rara vez desaparecía el dolor y cuando lo hacia era por breves periodos de tiempo (109.102). Estas caracteristicas se correspondían con el conocido síndrome de Dejerine-Roussy.
La observación animal permite ver dos tipos sistemas de conducción. Uno primitivo grueso, de representación bilateral y con estancia previa en el tronco del encéfalo que constituye el Paleoespinotalamico, que conduce casi exclusivamente sensaciones dolorosas y es propio de las aves . Otro sistema de conducción filogenéticamente de aparición posterior , que es el Neoespinotalamico , propio de mamíferos y mas aun de primates que recoge las proyecciones contralaterales de las vías sensitivas.
Por el sistema Paleoespinotalamnico, caminan las vías sensitivas que tras hacer estancia en el tronco del encefalo y mezclarse con el sistema reticular, pasan bilateralmente hacia ambos talamos, por dentro de la lamina medular interna y son conocidos como núcleos intralaminares de tálamo y forman lo que se conoce como vías extralemniscales de las sensibilidades (43,150,174,198,).
Esto demostraba que no todos los estímulos sensitivos pasan por los núcleos somatosensoriales de ambos talamos, sino que muchos de ellos eran extralemniscales , constituyendo las fibras retículo-talamicas de disposición medial al tracto espino talamico que terminan en los núcleos intralaminares del tálamo: Paracentral, Lateral Central, Centro Mediano y Parafascicular. Este sistema de transporte sensitivo es primitivo, transporta sensaciones profundas, dolorosas y mal localizadas . Mientras que las sensaciones que desembocan en los núcleos VPM y VPL también de ambos talamos son mas concretas superficiales y escasamente dolorosas .
De forma que la lesión de los núcleos VPL y VPM; solo rompía el equilibrio de los relés de las sensaciones no dolorosas y no conseguían accion terapéutica ( 8174,201,224).
Nauta demostró que las vías extralenniscales del dolor eran multipless tanto en animales como en el hombre y sus proyecciones colaterales terminaban en los núcleos intralaminares del talamo (184,185).
En los años 1960 se demostró por varios autores, que las lesiones de los núcleos intralaminares se seguían de la desaparición del dolor crónico
La lesión tenia que ser bilateral y simetrica y por dentro de la lamina medular interna y en ambos talamos para que fuera efectiva. Esto se había demostrado en necropsias).
El componente emocional del dolor desembocaba en el lóbulo prefrontal através de las proyecciones sensitivas que llegaban al núcleo DM, lo que permitía al Telencefalo el análisis de la sensación dolorosa Se sabia desde las celebres Leucotomias practicadas por el premio Nobel Egas Moniz en 1927, que se modificaba la personalidad del paciente que las recibía pero ademas aumentaba el umbral del dolor y era frecuente escuchar al paciente decir que el dolor ya no le importaba. Este mismo efecto se obtenia con las lesiones del Cingulo Rostral ,del nucleo DM y del nucleo anterior del talamo asi como lesiones practicadas en el lobulo prefrontal
De forma que las lesiones propuestas para analgesiar los dolores cronicos eran; CM, PF y DM, que se extendian a los núcleos reticulares que recibian el dolor extralemniscal y el DM, que vehiculaba el dolor a los lóbulos prefrontales informándoles del componente DM, que vehiculaba el dolor a los lóbulos prefrontales informándoles del componente doloroso, el sentimiento del dolor
Lesiones talámicas bilaterales
Material y Método.
Criolesion talamica bilateral
Desde los años comprendidos entre 1969 y 1980, se operaron 50 pacientes que sufrián dolor resistente a todos los procedimientos médicos y quirúrgicos conocidos.
La intervención se hacia colocando con anestesia local un marco estereotáxico diseñado por Leksell, que se colocaba en el craneo sujeto con tres fijadores que entraban en el cráneo hasta la tabla interna previa inyección de anestesia local
La localización de los núcleos se hacía previa ventriculografía a través de un agujero de trepano bilateral y simétrico a nivel de la sutura coronal y a 2,5 cm de la linea media. El contraste que se introducía en el sistema ventricular era una mezcla de aire y Dimer X. La cantidad total era de 8 cc. Era de capital importancia practicar la lesión en el agujero de Monro, para que al rellenar el III ventrículo y pudieramos ver las comisuras grises anterior y posterior (CA y CP) con claridad..
Los núcleos a lesionar se localizan inmediatamente por fuera del III ventriculo con la siguiente localización, extraídas del Atlas de Schatelbram y Bayley.
X.- 6mm por delante de la CP
Y.- 2mm por encima del plano CA-CP
Z.- Pared lateral externa de lII ventriculo.
Las lesiones se hacían con nitrógeno liquido que circulaba por dentro de una canula metalica de 2,4 mm de diametro y diseñada por Cooper. (Cooper Criosurgery Systen)
El volumen total de la lesión es un cilindro de 6 mm. de diámetro por 12 mm de generatriz.
La lesión se empezaba enfriando progresivamente la punta de la canula hasta llegar a 200º bajo cero.
Terminada la criotalamotomia, se cerraban las heridas craneales y se trasladaba el paciente a su habitación, totalmente despierto
Resultados según la Etiología del Dolor
Numero Resultado inmediatos Resultados tardios tiempo observacion
Tumoral
39 pacientes
39 Sin dolor 100%
36 sin dolor 92,3% 2 moderado 5.1% 1 igual 2.5%
Entre 1 y 6 meses 1 esta bien 30 años después Linfoma abdominal
Desaferentizacion
11 pacientes
11 sin dolor 100%s
2 sin dolor 18,18% 6 moderado 63.6% 3 igual 27,27 %
Entre 6 meses y 8 años
Complicaciones.
1 paciente con muy mal estado y metástasis múltiples fallecio 3 dias más tarde por hemorragia intraparenquimatosa
3 crisis convulsivas
1 sindrome de de Parinaud transitorio
31 paciente presentaron Sindrome frontal, indiferencia afectiva entre 2 y 7 semanas de duracion .
.
Esta tesis esta hecha en tiempos donde los métodos analgesicos eran escasos y los de utilización quirúrgica en la analgesia eran mutilante proporcionando en el postoperatorio mas desaferentizacion que analgesia.
Los resultados fueron mejores en los pacientes con dolor de origen tumoral, donde un 92,3 5 murió sin dolor. Muy posiblemente la menor duración de su vida después de la intervención fue responsable de estos resultados
En los pacientes con dolor de desaferentizacion los resultados fueron pobres ya que solo un 18,18% vivieron y murieron sin dolor. Estos pacientes vivieron mas tiempo y ello condiciona la recidiva del dolor.
Las complicaciones son escasas, ya que el paciente que murió tenia muy mal estado, como la mayoría de los pacientes tumorales operados.
El síndrome frontal que presentaron muchos pacientes, era beneficioso por que el dolor era entonces un hecho que dejaba de preocuparles y les permitía vivir con cierto bienestar.
Esta técnica en manos de un conocedor de la cirugía ESTEREOTÁCTICA tenían una mortalidad muy escasa y era muy efectiva el dolor es
Conclusiones
1.- La lesion de los núcleos intralaminaares del talamo con criotalamotomia produce un alto porcentaje de analegesia. Inmediata a la cirugía.
2.- Pasadas dos semnas los buenos resultados se reducen al 70%.
3.- En los enfermo portadores de neoplasia los resultados fueron superiores. La indiferencia del sindrome frontal les permitio vivir mas bondadosamente.
4.- Los pacientes con lesiones cutaneas, mejoraban de ellas marcadamente
5.- Un alto porcentaje de pacientes dejaron de ser adictos a los fármacos marcadamente , esto fue menor en los pacientes que sufrian dolores por desaferentizacion.
6.- En pacientes con dolores por desaferentizacion y que vivieron marcadamente mas tiempo después de la intervención los resultados fueron claramente peores. Los pacientes con disestesias previas a la intervenion rara vez mejoraron de ellas (neuralgias Posherpeticas, Arrancamiento plexo braquial).
7.- Se pueden considerar escasas las complicaciones y la dudosa mortalidad de esta estadistica es tambien baja
Creo que lo más interesante de este trabajo tardé tiempo en entenderlo.
Lo que yo buscaba era eliminar el dolor en el paciente , cosa que conseguía en un importante número de casos , pero lo que conseguí FUE QUITAR EL COMPONENTE SENTIMENTAL DEL DOLOR . AL PACIENTE YA NO LE IMPORTABA SU DOLOR AUNQUE SEGUÍA PERCIBIÉNDOLO . y esto era debido a que las vías que transportaban la sensación dolorosa a través del tálamo concretamente de sus núcleos intralaminares habían sido interrumpidas y los lóbulos frontales no se enteraban del dolor y por tanto ya no lo percibían como algo dramático .
Es decir las vías que transportan la sensación dolorosa para hacer concienciadas por el lóbulo frontal habían sido interrumpidas .
No habían desaparecido la señal dolorosa, es que no eran concienciadas por los lóbulos frontales, y desparecía el sentimiento del dolor.
PERSISTÍA LA EMOCIÓN, PERO LA LESIÓN A NIVEL DE LOS NÚCLEOS INTRALAMINARES, HABÍA ROTO LA VIA QUE LE PERMITÍA A LOS LÓBULOS FRONTALES, RECONOCERLO COMO SENTIMIENTO
LA EVIDENCIA DE QUE LOS NÚCLEOS CENTRO, MEDIANO, PARAFASCICULAR Y DORSO MEDIAL, ALOJADOS EN LA LAMINA MEDULAR TALAMICA, VEHICULAN LAS SEÑALES DOLOROSA HACIA LOS LOBULOS FRONTALES.
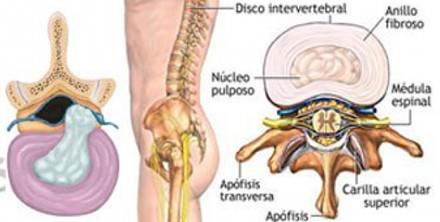
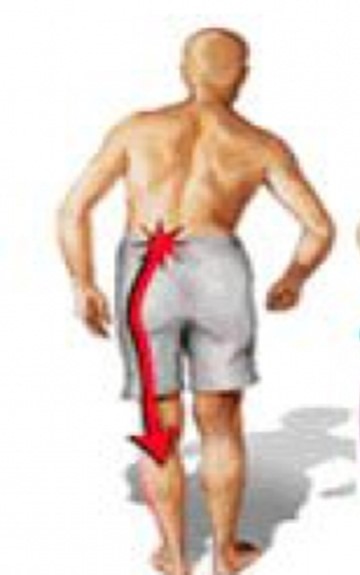
En los Servicios de Neurocirugía un 25% de todos los pacientes sufren patología raquídea sobre todo lumbar, de forma que esta muy tipificado el síndrome lumbo ciático de origen raquideo CR
Sin embargo, pese a que el ciático INTRARAQUIDEO, tienen unos 10 cm de longitud desde su origen medular lumbo-sacro hasta la salida por el agujero de conjunción, y el resto del ciático hasta el pie mide aproximadamente 1,15 cm. Excepcionalmente veíamos una ciática de origen EXTRARRAQUIDEO.
Mister y Bars, en 1934, operaran una hernia de disco lumbar con mucho exito, lo que hizo, que toda la patología ciática dolorosa, fuera interpretada de origen raquideo. Y desde entonces la ciática raquídea, predomina.
Hasta que en las últimas dos décadas, empieza a aparecer ciáticas, que se comportan de otra forma. Los dolores raquídeos ciáticos se apartaban de esta norma, y la cirugía en ellos era inefectivas, pese a que todos tenian osteofitos, solo entre un 5 y el 10%, eran debidos a compresiones de las raíces del ciático “ hernias de disco y estenosis del canal o de los agujeros de conjuncion”
De forma relativamente brusca y en el plazo de los últimos 10 años , el número de falsas ciáticas (FC), es decir que la patología no estaba en el raquis, se ha incrementado de manera considerable y sobre todo tengo la impresión de que esta verdadera epidemia de FC está ocurriendo de forma epidémica. No soy el primero que tiene esta impresión y además como coincide con la pandemia de coronavirus intentó relacionarlas y el único nexo que encuentro es que las personas que lo padecen o algunas de ellas , están mucho tiempo sentadas delante del televisor.
El síndrome de falsa ciática (FC): es una entidad conocida desde hace mucho tiempo , pero muy sometida a la confusión ya que la compresión del ciático a su paso bajo los glúteos y relacionado con el músculo piramidal pueden producir un dolor del nervio ciático que tiene una topografía muy similar a la ciática raquídea .
La única diferencia es que el signo de Lasegue (extension del miembro inferior, elongación del ciático) es negativo.
Como descripción general el dolor ciático de origen raquídeo (CR) se irradia hasta el pie de una forma concreta y por delante del miembro inferior por el contrario la falsa ciática nace en el glúteo y se irradia por la cara posterior del miembro inferior, es to como generalidad, porque en la practica no es fácil distinguir la topografía del dolor..
A este síndrome FC, se le ha llamado de muchas formas, aunque el origen de las molestias dolorosa, siempre es el mismo.
Empìeza en mitad del gluteo, ingle y cadera de manera suave y se irradia por la cara posterior y lateral del muslo de manera poco concreta y al llegar a la pierna el dolor suele ser aun mas difuso, aunque algún caso he visto con fuerte dolor e impotencia funcional del tobillo.
De manera genérica podemos decir que el FC es de suave comienzo y posterior y el CR, es muchas veces, mas concreto en su distribucuion y de lo localización anterior y sobre todo el signo de Lassegue es positivo.
No es infrecuente la aparicion de manera irregular de espasmos musculares a nivel de las pantorillas
Y un cierto grado de distimia, donde predomina la ansiedad
ANATOMÍA DEL CIÁTICO
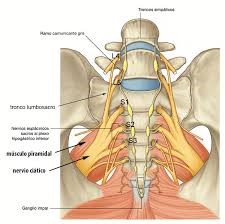
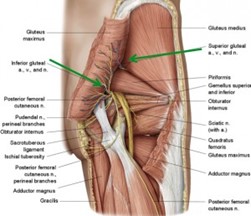
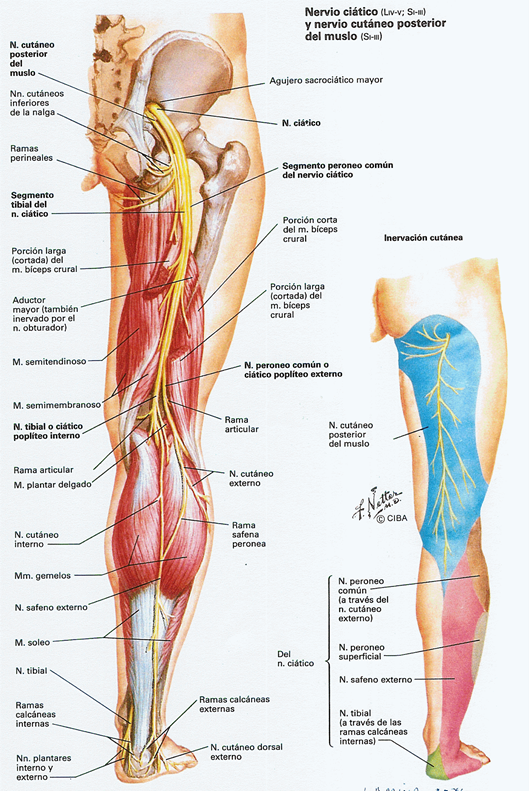
El nervio isquiático, más conocido como nervio ciático, es el nervio más voluminoso y largo del ser humano y otros mamíferos. Rama terminal del plexo sacro, es un nervio mixto que nace de las raíces de L4, L5, S1, S2 y S3. Comienza en la pelvis y sale de ella a través del foramen ciático mayor hacia la región glútea, bajo el músculo piriforme, sigue verticalmente a lo largo de la cara posterior del muslo donde termina bifurcándose en el nervio tibial y el nervio peroneo común. Fuera de la acción que pueda tener por sus ramos terminales, destinados a la pierna y al pie, el ciático es un nervio esencialmente motor, que asegura la flexión de la cadera
sobre el muslo
Nace del cuarto y quinto nervios del plexo lumbar a través del tronco lumbosacro, y de la primera, segunda y tercera raíces saacras. La convergencia de estas raíces en un solo tronco se efectúa en la escotadura ciática mayor, en el borde inferior del músculo piramidal y por detrás del borde posterior del obturador interno, por debajo de la espina ciática.
Luego de su origen, el nervio sale de la pelvis por la parte inferior de la escotadura ciática mayor, por donde penetra en la nalga. Inclinándose entonces de arriba abajo y de medial a lateral, desciende verticalmente por un profundo canal que le forman el isquion medialmente y el trocánter mayor lateralmente, detrás de los músculos pelvitrocantéricos. A su salida de este canal penetra debajo de la porción larga del bíceps femoral y corre enseguida a lo largo de la cara posterior del muslo hasta el vértice de la fosa poplítea, en donde termina bifurcándose.
En el foramen ciático mayor, ocupa la parte lateral del espacio infrapiriforme. Es lateral a los vasos pudendos internos y glúteos inferiores y a los nervios pudendos, al obturador interno y anal. El cutáneo femoral posterior está situado por detrás de él.1
En la región glútea, desciende en un tejido celulo adiposo que separa al glúteo mayor detrás del plano de los gemelos, del obturador interno y del cuadrado femoral adelante, los que los separan arriba de la cara posterior de la articulación de la cadera. Recibe aquí una rama de la arteria glútea inferior: la arteria del nervio ciático fina y larga. Esta arteria y el nervio cutáneo femoral posterior están situados por detrás de él y describen un trayecto paralelo.
En la parte posterior del muslo, se aplica sobre la cara posterior del aductor mayor, después sobre la porción corta del bíceps femoral. El nervio cutáneo femoral posterior lo abandona para descubrir un trayecto más superficial. En la parte superior del muslo, el ciático es lateral al tendón de la cabeza larga del bíceps y cubierto por las fibras inferiores del glúteo mayor y más abajo de la cabeza larga del bíceps lo cruza de medial a lateral. El nervio se sitúa en el intersticio formado por el bíceps, lateralmente, y el semimembranoso, medialmente. El bíceps cubre el trayecto del nervio hasta su inserción en la cabeza del peroné. En todo este trayecto queda en relación con la arteria del nervio ciático y las anastomosis de las ramas perforantes de la arteria femoral profunda, acompañadas por las anastomosis venosas correspondientes.
El nervio ciático suministra siete ramos colaterales destinados a los músculos de la región posterior del muslo y a la articulación de la rodilla. Estos ramos colaterales son el nervio superior del músculo semitendinoso, el nervio de la cabeza larga del músculo bíceps femoral, el nervio inferior del músculo semitendinoso, el nervio del músculo semimembranoso, el nervio del músculo aductor mayor, el nervio de la cabeza corta del músculo bíceps femoral y el ramo articular de la rodilla. Excepto los dos últimos, estos ramos nacen generalmente en la parte superior del muslo, bien de forma aislada bien por medio de uno o dos troncos comunes:
1. El nervio superior del músculo semitendinoso penetra en el tendón común de los músculos bíceps femoral y semitendinoso e inerva al músculo semitendinoso superiormente a la intersección tendinosa.
2. El nervio de la cabeza larga del músculo bíceps femoral es oblicuo inferior y lateralmente, y penetra en el músculo por la porción media de su cara anterior.
3. El nervio inferior del músculo semitendinoso penetra en la porción media del músculo.
4. El nervio del músculo semimembranoso comprende uno o dos ramos que abordan el músculo por su cara lateral.
5. El nervio del músculo aductor mayor nace bien del nervio ciático bien del nervio del músculo semimembranoso, y se distribuye por la porción medial del músculo aductor mayor.
6. El nervio de la cabeza corta del músculo bíceps femoral se separa del nervio ciático a la altura de la porción media del muslo y se distribuye en la parte media del músculo.
7. El ramo articular de la rodilla nace de la cara anterior del nervio ciático o del nervio de la cabeza corta del músculo bíceps femoral. Termina en la porción lateral de la articulación de la rodilla.
Denominaciones de este sindrome
Se le llama de varias formas, ya que sobre una anatomía constante, diversas causas lastiman al ciático:
Describire solo dos:
Bursitis trocantérea La bursitis trocantérea, o trocanteritis, como tal, es la inflamación de las bolsas serosas que se sitúan en la extremidad proximal del fémur. Debido a que dichas bolsas están rellenas de líquido sinovial, están expuestas a todos aquellos procesos de índole inflamatoria, que afectan a la cadera, como, por ejemplo, la artritis reumatoide. Sin embargo, la mayoría de las trocanteritis están causadas por microtraumatismos de repetición y la inflamación consiguiente que lo perpetua.
HIPERTROFIA DEL PIRIFORME
El músculo piriforme participa en casi todos los movimientos que realiza con la parte inferior del cuerpo, desde caminar hasta cambiar el peso de un pie al otro. Por debajo del músculo se encuentra el nervio ciático. Este nervio va de la parte inferior de su columna vertebral completamente hacia abajo en la parte trasera de su pierna hasta su pie.
Los traumatismos en la zona
Son muy frecuentes en las caídas sobre los glúteos, a veces anodina, pero que pasado poco tiempo empieza una clínica, con dolor ciatico
Lesionar o irritar el músculo piriforme puede causar espasmos musculares. El músculo también podría inflamarse o tensarse a causa de los espasmos. Esto pone presión en el nervio que pasa por debajo, causando dolor. Sentarse durante períodos prolongados
Hacer demasiado ejercicio
Correr, caminar o hacer otras actividades repetitivas
Practicar deportes
Subir escaleras
Levantar objetos pesados.
Otras varias causas, como Síndrome de los glúteos medios, son tenidas en cuenta.
Pero patología inherente a la inflamación del nervio y produccion de neuritis, solo de manera excepcional son mencionadas.
Lo que si es cierto que el nevio ingresa en el miembro inferior después de su nacimiento, por la escotadura que se produce entre el musculo Piramidal y los gemios
Incidencia El proceso puede afectar a ambos sexos y a todas las edades, pero existe un marcado predominio en las mujeres (3:1 frente a los varones), que suelen ser obesas y encontrarse entre los 40 y los 60 años de edad3. En el ámbito deportivo, se trata de una afección muy frecuente en los porteros de fútbol y en aquellos deportistas que sufren, como consecuencia de su actividad deportiva, un excesivo rozamiento de la región, como ocurre, por ejemplo, en los corredores de maratón.
Es claramente mas frecuentes en mujeres Y CUANDO ESTAN EMBARAZADAS.
Durante las maniobras del coito, donde las mujeres adoptan una postura de abducion de los miembros inferiores, el que las mujeres sientan dolor glúteo intenso y pasajero, es muy frecuente.
Las personas que hacen trabajos rudos no suelen padecerla y si los que esta mucho tiempo sentados
Algunos pacientes con FC, han sufrido infecciones, próstata, vejiga y además tienen brotes de dolores poliarticulares.
Como si estuviéramos ante una enfermedad mixta, donde la compresión del ciatico en los glúteos, necesitara en ocasiones de una enfermedad inflamatoria previa.”neurodegeneracion”
Una serie de maniobras empeoran o mejoran este síndrome:
FLEXIÓN, EXTENSIÓN Y SEMIFLEXIÓN DE LA CADERA
La relación de la región lumbar con los miembros inferiores modifica el dolor
Las posturas, lumbo-muslo , elongación, flexión y semiextension, modifican de manera importante el dolor y lo agravan o mejoran, e incluso lo localiza de forma diferente.
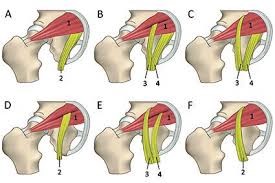
Una serie de dibujos, muestran como el angulo de flexión de la cadera modifican el trayecto ciático y lo doblan, lo que les desencadenan o mejoran el dolor.
Estas maniobras son espontaneas y no estudiadas y los pacientes que la sufren al estar sentados, al disminuir el angulo del ciático en la cadera.
Una serie de esquemas, intentan demostrar el angulo del muslo con el tronco, como intervenientes en esta patología
La curva del ciático, es nula y por tanto al pasar por el desfiladero del musculo piramidal, no se comprime. Pese a ello la bipedestación es dolorosa, y el caminar la empeora.
Es la peor de las posturas, el ciatico se dobla 90º
Postura que frecuentemente adoptan los pacientes con FC y que a ratos los mejora.
Tratamiento de esta entidad.
Tienen un tratamiento no ordenado.
Analgésico y antinflamatorios, confortan.
Relajantes musculares para dormir.
La rehabilitación depende de los autores. Algunos le prestan mucho valor y utilidad, y otros la desestiman por inútil.
Conclusiones.
La Falsa ciática, se debe a un fenómeno compresivo en el desfiladero del ciático por los músculos piramidales y gemius.
Su origen es traumático en ocasiones
Otras espontáneos
Es muy posible que forme parte de las enfermedades neurodegenerativas.
La epidemia del coronavirus puede ser un factor desencadenante por dos razones
Porque agrega inflamación al ciático
Por estar mucho tiempo sentado.
Su duracion es irregular asi como su intensidad.
Desde un trauma, van apareciendo molestias fisicas y psíquicas de manera irregular.
Evoluciona por brotes y a veces termina con cierta brusquedad
Bibliografía
Rouviére H, Delmas A (2005). Anatomía humana descriptiva topográfica y funcional. Barcelona: Masson.
Testut, L., Latarjet, A. Tratado de anatomía humana. Barcelona: Salvat. 1984.
Latarjet, M., Ruiz Liard, A. Anatomía humana. Médica Panamericana; Buenos Aires, Argentina 2014.
Anatomía Humana. Tomo 3. Miembros, 11va Edición – Henri Rouvière-
-Martínez F, Valverde A (2006). Anatomía de los plexos lumbar y sacro En: Aliaga L, Anestesia Regional Hoy, (pp 84-263) 3ª ed. Barcelona. Permanyer.
-Netter FH (2003). Atlas de anatomía humana. Barcelona: Masson.
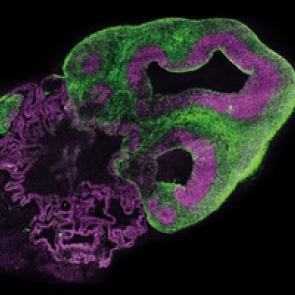
Mini cerebro humano y de gorila S.Benito-Kwiecinski/MRC LMB/Cell
Imagen de un organoide cerebral obtenida mediante microscopía de fluorescencia. En verde, se observan las neuronas, y en magenta, las células progenitoras. [Lancaster et al./ Nature]
El cerebro humano es una de las características más distintivas de nuestra especie. De todos los primates, somos los que han logrado un órgano de mayor tamaño. Tanto así, que nuestro cerebro triplica al de un gorila o un chimpancé, nuestros parientes más cercanos.
Se ha intentado entender la causa de esta rápida expansión evolutiva con estudios comparativos con animales como ratones o macacos. Pero la investigación se enfrentaba al gran problema de abordar las etapas más tempranas del desarrollo cerebral, antes incluso de la formación de las primeras neuronas, periodos en los que se sospechaba que se hallaba la clave del mayor tamaño del cerebro humano.
La evolución hizo que el cerebro humano aumentara de tamaño respecto al de sus antecesores por un mecanismo hasta ahora desconocido. Gracias al estudio en el laboratorio de organoides cerebrales, se ha podido hallar que la clave del misterio está en el prolongado tiempo de transformación de las células que dan lugar a las neuronas. Que este paso sea más largo permite que surjan más células progenitoras y por tanto, más neuronas en un futuro.
Patrones de expresión genética específicos en humanos, en comparación con otros grandes simios, se encuentran detrás del fenómeno.
Los organoides son cultivos celulares tridimensionales, inicialmente de origen epitelial, generados por células stem/troncales (adultas, embrionales o inducidas) que reproducen parcialmente las características de sus tejidos de origen. Constituyen un sistema más apropiado que las clásicas líneas celulares de crecimiento bidimensional en plástico para estudios de biología del desarrollo y celular e histología, así como de procesos de tumorogénesis y de medicina regenerativa.
Con la combinación adecuada de nutrientes, las células madres humanas derivadas de la piel pueden agregarse de forma espontánea y convertirse en fragmentos de tejido cerebral. Así lo demuestra un estudio recién publicado en Nature.
Crear un cerebro artificial completo tal vez tarde aún varios años, pero los organoides cerebrales, del tamaño de guisantes, desarrollados en este trabajo podrían resultar de utilidad en la investigación de enfermedades neurológicas humanas.
Con anterioridad ya se habían utilizado células madre humanas para crear estructuras parecidas al ojo e incluso tejidos similares a la corteza cerebral. Sin embargo, en este último estudio, los científicos obtuvieron fragmentos de tejido neural más grandes y más complejos. Primero cultivaron las células madre en un gel sintético parecido a los tejidos conectivos naturales que se hallan en el cerebro y en otras partes del cuerpo. A continuación, introdujeron los grupos incipientes de células en un biorreactor para proporcionar al tejido nutrientes y oxígeno. Para sorpresa de los investigadores, las manchas crecieron hasta parecerse a cerebros de fetos en la novena semana de gestación.
Bajo el microscopio, las distintas regiones cerebrales parecían interaccionar unas con la otras. Pero entre las muestras de tejido no se formaba ninguna estructura fisiológica reconocible. Las estructuras conseguidas, no eran equiparables a un cerebro. Los organoides carecían de vasos sanguíneos, lo que podría explicar su reducido tamaño, de unos 3 o 4 milímetros de diámetro como máximo, incluso después de cultivarlos durante 10 meses o más. La maduración de este órgano en el embrión probablemente esté guiada por señales de crecimiento procedentes de otras partes del cuerpo
A pesar de estas limitaciones, los autores utilizaron el sistema para modelar los aspectos clave de la microcefalia, una enfermedad que causa un crecimiento muy deficiente del cerebro y deterioro cognitivo. El estudio del desarrollo neurológico no reprodujeron en roedores características que se investigaba para la formación del cerebro.
Pero cuando los organoides se crearon desde células madre de la piel de un humano con microcefalia, estas no alcanzaban el tamaño de los obtenidos a partir de células madre de una persona sana. Se deducia que la diferenciación era prematura en las células madre neurales dentro de los fragmentos de tejido microcefálico, y que se debía a que las células progenitoras para el crecimiento normal del cerebro, se habían agotado
Se elaboro la hipótesis de que hacían falta la inmersión en los cultivos de otros tejidos no neurales para reproducir la microcefalia.
En la actualidad los organoides cerebrales, también conocidos como mini cerebros, son una nueva biotecnología con el potencial para cambiar nuestro entendimiento sobre el desarrollo y las enfermedades neurológicas.
El cerebro humano presenta una estructura tridimensional que los organoides diseñados en el laboratorio pueden en parte reproducir Muchos de estos experimentos se están llevando a cabo en la Estación Espacial Internacional (ISS ), pero uno de los más interesantes es el que está realizando el equipo del biólogo Alysson Muotri, de la Universidad de California en San Diego, quien trabaja con un grupos de células nerviosas llamadas mini-cerebros en la ISS Los organoides se cultivaron a partir de células madre en el laboratorio, antes de ser colocados en una caja y enviados al espacio, donde Muotri señala que probablemente «se están reproduciendo a un ritmo acelerado”.
Lo desafiante es que estos organoides emiten ondas cerebrales, patrones complejos de actividad neuronal, similares a los de los bebés prematuros.
Este impresionante descubrimiento, ponen de manifiesto que los mini-órganos cultivados tienen funciones cerabrales, aunque primitivas.
Posteriormente Muotri, conecto los mini cerebros a robots para leer su actividad neuronal. Estos hallazgos podrían expresar que se esta acercando a la producción de vida acercando a la capacidad de generar al menos una vida parcialmente consciente en el laboratorio, un desarrollo que durante mucho tiempo ha sido poco más que una fuente para relatos de ciencia ficción.
Los mini cerebros: son conjuntos de neuronas comparativamente simples que simulan algunas características de los cerebros completos, pero esto es solo una ilusión, pero el hallazgo, sugiere que los mini cerebros podrían ser aún más complejos de lo que se creía anteriormente. Si estas ondas cerebrales son una señal de que los organoides podrían ser conscientes, los neurocientíficos necesitarán lidiar con un importante dilema ético, ya que la experimentación continua podría significar que se manipula la vida humana y consciente de sí misma. Los mini cerebros, en la actualidad se desarrollan a partir de células madre que acaban dando lugar a pequeñas bolas de 4 milímetros de tamaño. Este diseño tridimensional del tejido neural, lo convierte en el modelo más realista del cerebro que tenemos hasta la fecha. compuestas por hasta 3 millones de células (neuronas y células gliales). En 2013, el trabajo con mini cerebros, mostró, que sus células son capaces de organizarse solas en estructuras que recuerdan al cerebro De esta forma, imitan las dinámicas espacio temporales de la neurogénesis (formación de neuronas) y el desarrollo de circuitos. Durante su desarrollo las neuronas ‘migran’ para dar lugar a distintas capas y conexiones (Bob Jacobs, Colorado College) Estos modelos polemizan por la incertidumbre que genera y, sobre todo, porque toca algo que nos define como humanos: la conciencia. “Se piensa que el soporte físico que la genera son los patrones eléctricos y las conexiones entre neuronas. Por lo tanto, científicamente hablando, la conciencia puede estar en el cerebro. Si estamos creando estructuras similares a este órgano en un momento dado no puedan tener conciencia. No quiere decir que esto sea posible pero sí probable.
Estos organoides comenzaron a producir patrones eléctricos como los generados por un cerebro normal.
Desde aquí a que los organoides permitan el diseño de cerebros con sentimientos, y todo el contexto que lleva consigo, es demasiado imaginar Imaginar que los aspectos éticos se adelanten a lo somático, nos confunden, aunque se trate solo de un experimento.
Pero no vamos a frenar la ciencia por no saber lo que pueda ocurrir, o porque exista la posibilidad de que en un futuro se desarrolle conciencia en los mini cerebros. Como afirma Santaló, nunca averiguaremos estas incertidumbres si no las investigamos.“Debemos actuar con cautela y a cada paso dado, parar y reflexionar”. A partir de aquí, infinidad de ideas se van a desarrollar Un ejemplo es el que sigue: Crean un mini cerebro capaz de generar movimiento muscular Escrito por Álvaro Justo, 4 de abril de 2019 a las 17:30
Un grupo de científicos de la Universidad de Cambridge ha conseguido replicar en miniatura un cerebro embrionario con el fin de estudiar su desarrollo.
Han desarrollado un mini cerebro capaz de generar movimiento muscular. Se trata de un organoide del tamaño de una lenteja, obtenido a partir de células madre humanas. Los resultados se han publicado en la revista Nature Neuroscience.
Representación del cerebro humano Los organoides cerebrales se utilizan en el mundo científico para estudiar el crecimiento del cerebro, ya que, al estar constituidos por tejido neuronal generado a partir de células madre, forman estructuras similares a las de los cerebros embrionarios humanos. Para poder observar su desarrollo lo que se hace es sumergirlos en un líquido con alto contenido en nutrientes. El inconveniente que tienen es que, al carecer de vasos sanguíneos, no pueden acceder al oxígeno y los nutrientes necesarios para sobrevivir mucho tiempo. El equipo de investigación de Cambridge ha dado con un método para solventar esta barrera. Consiste en crear una estructura de membrana porosa para que el organoide crezca en un entorno aire-líquido que permite un mejor acceso a los nutrientes y al oxígeno. Esto facilita un periodo de vida más largo de la estructura, lo que se traduce en estados de desarrollo neuronal más avanzados. Gracias a ello, los científicos han podido observar en el mini cerebro una conectividad neuronal similar a la existente en un embrión humano de entre 12 y 16 semanas. Para comprobar la capacidad funcional del mini cerebro, los científicos colocaron junto a él un pedazo de médula espinal de un embrión de ratón con el músculo adyacente de la espalda. El resultado mostro que los axones de algunas neuronas específicas crecieron de forma espontánea para conectarse con la médula espinal, llegando a contraer el músculo con las señales eléctricas emitidas.
Si esto progresa, la manipulacion de cultivos de células nerviosas creadas en el laboratorio, tendrían una indicación enorme para reparar patologías relacionadas con el sistema nervioso. El análisis del nacimiento y el desarrollo de las redes de conectividad neuronal puede ayudar a comprender el funcionamiento de patologías como la epilepsia, la esquizofrenia, el autismo, la depresión o los distintos tipos de demencias. La ciencia nos muestra el camino para mejorar las condiciones de sus vidas.
Bob Jacobs en Colorado College ha dado un paso sorprendente. Durante su desarrollo las neuronas ‘migran’ para dar lugar a distintas capas y conexiones.
Esto como siempre en biologica, tienen un camino, muy difícil de imaginar, pero hasta ahora es sorprendente
La investigación se enfrentaba al problema de abordar las etapas más tempranas del desarrollo cerebral
“La hipótesis en la que todo el mundo pensaba que tendría que haber algún mecanismo que aumentara la proliferación de las células progenitoras antes de crear neuronas”. segun piensa Silvia Benito Kwiecinski, del laboratorio de Biología Molecular del Consejo de Investigación Médica del Reino Unido.
El equipo detrás de la nueva investigación, dirigido por la bióloga Madeline Lancaster, tenía un as bajo la manga para superar las dificultades: los organoides cerebrales. Lancaster formó parte del equipo que creó los primeros mini cerebros cultivados en laboratorio en el 2013.
Los organoides cerebrales suponen las réplicas del cerebro más realistas que tenemos hasta la fecha. Al tener la capacidad de autoorganizar sus células –neuronas y células gliales–, proveen a los investigadores de una herramienta muy útil para observar cómo se desarrolla este órgano.
El grupo de Lancaster empleó mini cerebros derivados de células humanas, de gorilas y de chimpancés para estudiar qué ocurría antes de que aparecieran las neuronas y qué diferencias presentaban los tres modelos entre sí.
Benito, se centró en observar el comportamiento de las células progenitoras de neuronas muy pronto en el desarrollo embrionario, unas cuatro semanas tras la concepción.
Hubo un proceso en concreto que llamó su atención: la transformación de las denominadas células neuroepiteliales a células gliales radiales. Entre ambas progenitoras se daba un proceso de transición con células intermedias.
“Lo que se sabía gracias a estudios de ratones era que este cambio entre neuroepiteliales y gliales era algo que ocurría muy rápido. Nunca se había visto si había algo intermedio. Vieron que hay otro proceso ocurriendo en los cerebros de los simios”, .
En el caso de los organoides humanos, este periodo de transición tardaba más en comenzar e iba ligado a ciclos celulares más cortos en las células neuroepiteliales. Es decir, se multiplicaban más rápido por lo que luego se desarrollarían más neuronas. Por eso, el equipo observó cómo los mini cerebros humanos crecieron más que los de los simios
Detrás de la diferencia en esta especie de metamorfosis celular se encontraba un gen, el ZEB2. Este se activó antes en los organoides del cerebro de los gorilas que en los organoides humanos, algo que pudieron determinar mediante secuenciación del ARN.
“Este gen era uno de los principales candidatos. Es un gen que regula la transición epitelio-mesénquima. Es decir, hace que células epiteliales se hagan menos epiteliales. Y eso es justo lo que ocurre en el cambio de neuroepilelial a glial”.
Los investigadores señalan que los organoides son un modelo, que aunque sea el mejor hasta la fecha, no replican a la perfección los cerebros reales.
Referencias
Yin, X., Mead, B. E., Safaee, H., Langer, R., Karp, J. M., & Levy, O. (2016). Engineering Stem Cell Organoids. Cell stem cell, 18(1), 25–38. doi:10.1016/j.stem.2015.12.005 Grebenyuk, S., & Ranga, A. (2019). Engineering Organoid Vascularization. Frontiers in bioengineering and biotechnology, 7, 39. 10.3389/fbioe.2019.00039https://stemcellsjournals.onlinelibrary.wiley.com/doi/full/10.1002/stem.1290
LEYRE FLAMARIQUE, MADRID 28/10/2019 06:00Actualizado a28/10/2019 09:26
No sé si nos damos cuenta, pero estamos en medio de una batalla, creciente entre mujeres y hombres.
Esta batalla, pero en mínimos lleva mucho tiempo, pero se ha activado y de manera creciente, en los últimos tiempos.
Las manifestaciones continuas y tumultuosas y las peticiones de la mujer son crecientes.
De entrada, digo, que estoy con ellas, que han sido vejadas durante la historia y continuan siéndolo y esto tiene que acabar.
Pero ojo, lo que ha de pasar no esta escrito.
Si la mujer gana la batalla y es muy posible que así sea. El hombre no solo no va a mandar, sino que “no puede tener hijos y la mujer si”. Incluso con una célula de su piel modificada, se puede fecundar sus óvulos y todo queda en casa.
De hecho en el reino animal la autofecundacion esta ampliamente comprendida
Animales hermafroditas
No le pasa lo mismo al hombre, no tiene donde depositar sus espermas, salvo que fabrique un útero.
Pero hasta ahora, la cosa no va así.
Si el hombre no tiene una mujer a la que fecundar, no proliferara la especie. La homosexualidad tan creciente y ganando terreno, no tiene esto solucionado.
La mujer lo tiene y de sobrado.
Ya hace un siglo, Aldoux Husley, escribió Un Mundo Feliz, donde proponía un sistema, pera crear hominidos de laboratorio perfectamente adoctrinados y drogados contra la ansiedad, que terminaba con la diferencia hombre mujer.
Todos iban por el mismo camino y estaban en el laboratorio preparados, para mandar o servir En el centro de incubación y condicionamiento de la central de Londres trabajaban 300 fecundadores y donde se unía un óvulo y un espermatozoide en un tubo de ensayo y se multiplicaban dando lugar de 896 embriones idénticos este proceso se realizaba paradójicamente deteniendo el crecimiento normal era el principio de producción en masa aplicado a la biología
Los embriones se mandaban a la sala de predestinación social para crear seres humanos socializados con un futuro determinado se les controlaba el nivel de oxígeno según la casta a la que debían pertenecer se los proporcionaba diferente cantidad de oxígeno para que fueran más o menos inteligentes.
Los épsilon eran la casta más baja los alfas la más alta independientemente de su pertenencia todos estaban educados para amar su destino .
Todos iguales, el cerebro bien lavado y una jerarquía biológica inmutable.
Esto hoy con los procedimiento que tenemos, seria fácilmente conseguible
Pero la lucha no tiene por qué colocar al varón como jefe, lo puede ser una mujer perfectamente.
De formas que de aquí van han surgir cosas. Ni siquiera hace falta esta industria tan compleja
La primera el varón ya no es necesario para la evolución. No tiene a quien preñar
La mujer es imprescindible.
Como esto va a ocurrir y que va a proporcionar.
¿Va ha desaparecer el hombre como animal, o convertirse en algo subrogado, cuando ahora es un rey?
No hay ni un solo día, en que un hombre no mate a una mujer o la destruya de alguna manera. Esto ya nadie lo aguanta y las mujeres menos.
Esto a nivel social, no se soporta e incluso muchos hombres, las apoyan.
De forma que podemos tener un futuro lleno de mujeres. “Que manden por su numero y por su cerebro mas lógico”.
E incluso como ellas tienen los materiales, para la gestación y fecundación, podrían determinar que solo fueran mujeres la descendencia. Es cuestión de quimica
El cerebro de las mujeres es mas pequeño, como los ordenadores, gasta menos y tienen una infinita capacidad y ademas tienen dos cromosomas XX. Y no para que uno sustituya al otro, sino con funciones que el hombre no tiene, sobre todo en lo referente a la visión y otras muchas funciones por descubrir.
El hombre tienen dos cromosomas X e Y. y esto no se si iguala a las capacidades de la mujer.
De forma que este libro tiene como objetivo alarmar al hombre, de que podemos desaparecer como especie, o la de no pertenecer a la especie dominante.
Esto va a pasar en milenios, y en el intervalo puede ocurrir cualquier cosa.
Y como el hombre todo lo ha solucionado, o muchas veces lo hace con la guerra, no se si este método va a valerle en el futuro, con alguien con libertad, capacidad de autorreproducirse y con otros objetivos no belicistas.
La llamada “ideología de género” afirma, alegremente, que la cualidad de femenino o masculino en el ser humano no viene dada por la naturaleza sino por la cultura; que no se nace hombre o mujer, sino que cada cual elige su sexo entre una multiplicidad según las propias inclinaciones o deseos.
Esto puede ser cierto, pero tenemos que aceptar las consecuencias.
Nunca en las especies, ha habido una guerra por el poder entre los géneros de los animales, ni tan siquiera cuando se convirtió en hominido.
El dominio lo tuvo casi siempre uno de ellos, el hombre en general, y los demás sumisos a aguantar.
De pronto las mujeres han decidido que esto se acabó, que no necesitan un mando masculino para seguir viviendo como especie, y “ han empezado una guerra”, sobre todo con inteligencia y manifestaciones, que dan tanto fruto. Se las pueden apañar solas
La ideología actual no lucha por el respeto a los homosexuales o “trans”, sino por la confrontación entre el hombre y la mujer.
La ONU, impone, universalmente, el feminismo radical, esto viene ya de lejos. no es algo moderno, Friedrich Engels, seguidores de Carlos Marx. Marx parten del capital y del trabajo, para lograr la revolución social y el poder. Engels teorizaba sobre la confrontación del hombre y la mujer, para lograr la desaparición de la familia. Esta ideología y adocenamiento machaca, incluso a los niños pequeños en las escuelas, y por la imposición de silencio. Ha obedecer se ha dicho. Como si esto fuera aceptado eternamente
Pero se dejan atrás, que el otro también pinta, y además a medida que se pisa mas a las mujeres, las están invitando a revelarse y empezar su lucha.
Y claro que lo han hecho. Pongan la TV un momento y lo verán día si y día tambien.
El sexo no reproductivo, movió al hombre de tal manera que empezó a utilizarlo de todas maneras con el fin de mantener quieta su ansiedad, tan en progreso y llego totalmente a desvincularlos.
Tenemos dos formas de usar el sexo.
Una para procrear y proliferar la especie.
La otra como órgano de placer.
La primera no es simplemente necesaria. Determinadas razas son muy prolíficas, y sustentan con creses la natalidad, mientras que muchos grupos eligen el sexo como placer, probablemente como droga, les es mas gratificante y caiga quien caiga.
Y aquí estamos.
Esto no es baladí, de aquí parten consecuencia y de ellas y repito, el hombre sale perdiendo, ya no cumple los parámetros de los seres vivos “la reproducción”.
Siempre le queda la oportunidad de actuar como un virus y comportarse como parasito. Pero esto no ha sido nunca nuestro sueño.
Mandar, mandar y volver a mandar y caiga quien caiga.
Todo el mundo se ha asustado con la noticia de los efectos negativos que han aparecido en un bajo porcentaje de los vacunados contra el Covid 19, con la vacuna de AstraZeneca . Las trombosis venosas, sobre todo de una entidad temible y difícil de diagnosticar, las trombosis de los senos venosos cerebrales.
La trombosis venosas, de uno de los varios senos del cerebro, para vaciar, la sangre desoxigenada al corazon, produce unos síntomas claros, que se deben a “entra la sangre por las arterias, al cerebro y no sale para el corazón por las venas”. Cuando se obstruye uno o varios SENOS, se produce una hichazon del, cerebro y después todas las consecuencias de la alteración de la circulación, aparecen en cadena.
Es decir la sangre entra en cerebro y con gran presión pero no sale.
El síntoma dominante es DOLOR DE CABEZA INTENSO
La trombosis puede aparecer en cualquier parte de nuestra biología, en los senos cerebrales o en las venas de los miembros inferiores
Estos son los síntomas de la trombosis de senos venosos cerebrales detectada en varios vacunados con la AstraZeneca
La trombosis del seno venoso cerebral (CVST) es la presencia de trombosis (coágulo sanguíneo) en el seno venoso dural, que recibe sangre de venas externas e internas del cerebro.
Los síntomas de esta enfermedad pueden incluir dolor de cabeza, visión anormal, cualquiera de los síntomas de un accidente cerebrovascular (debilitamiento de los músculos de la cara y extremidades de una mitad del cuerpo) e infartos.
El tratamiento se realiza con anticoagulantes, que suprime los coágulos sanguíneos), y raramente por trombolisis (destrucción enzimática del coágulo). En caso de que exista una causa oculta de la enfermedad, se realizarán exámenes para determinar la misma. La enfermedad puede agravarse por la acción de la presión intracraneal, que puede implicar una intervención quirúrgica, normalmente por shunt1 cerebral o derivación ventrículo peritoneal.
Nueve de cada diez personas con trombosis del seno venoso padecen dolor de cabeza, que tiende a empeorar con el paso de varios días, pero también puede desarrollarse espontáneamente un dolor de cabeza intenso.1 La cefálea puede ser el único síntoma de la trombosis del seno venoso cerebral.2 Varios pacientes también tienen síntomas propios de ACV: incapacidad para mover una o más extremidades, debilidad en una mitad de la cara o afasia. Esto no afecta necesariamente a una parte del cuerpo como en los accidentes cerebrovasculares más comunes.1
Son más comunes en mujeres y es conocido que desarrollan trombosis en el seno venoso cerebral después del parto.3 Estos ACVs afectan a una parte del cuerpo, normalmente a una mitad, pero ocasionalmente los ACVs son generalizados y raramente conducen al status epilepticus (crisis epiléptica única o repetidas).1
Si la presión intracraneal está muy elevado, el nivel de conciencia decrece, la presión sanguínea aumenta, se produce una bradicardia y el paciente adopta una pose anormal a su habitual.1
Porque se producen estas trombosis.
El 85% de los pacientes de esta enfermedad tiene al menos uno de los siguientes factores de riesgo:1
Su puede diagnosticar esta enfermedad con base en los síntomas ya mencionados; por ejemplo, la combinación de dolor de cabeza, signos de elevada presión intracraneal y anomalías neurológicas focales, o cuando causas derivadas del dolor de cabeza y anormalidades neurológicas, como la hemorragia subaracnoidea, han sido excluidas.1
Las pruebas de imagen son definitivas, para el diagnostico de estos procesos.
Hay varias formas de neuroescaneo que pueden determinar la presencia de la trombosis del seno venoso cerebral. Los edemas cerebrales e infartos venosos pueden hacerse visibles en cualquier modalidad, pero para la detección del trombo, el tipo de escaneo más usado comúnmente es la tomografía axial computarizada (CT) y la resonancia magnética (MRI). Ambas usan varios medios de radiocontraste para elaborar un venograma y visualizar las venas en torno al cerebro.1
TROMBOSIS SENO LONGITUDINAL SUPERIOR
La tomografía axial computarizada tiene una capacidad de detección que para algunos expertos supera la de la resonancia magnética. Esta prueba se basa en la inserción, a través de una inyección en la vena (normalmente en el brazo), de una sustancia radiopaca. Tras un tiempo, el que tarda el torrente sanguíneo en llevar la sustancia al cerebro, el escaneo se realiza. La sensibilidad de este test es de 75-100% (detecta de 75 a 100% de todos los coágulos presentes), y una especificidad de 81-100% (sería incorrectamente positivo en un 0-19%).5
La resonancia magnética venosa MRI , que emplea los mismos principios, pero la modalidad tiene la ventaja de ser mejor a la hora de detectar peligro para el cerebro como resultado del incremento de la presión en las venas obstruidas. No obstante, este mecanismo no se encuentra disponible en muchos hospitales y su interpretación puede resultar dificultosa.5
La analítica es muy eficaz para el diagnostico
Dímero-D
Un estudio de 2004 sugiere que el test sanguíneo Dímero-D, ya en uso para diagnósticos de otras formas de trombosis, se comportaba anormalmente (por encima de 500 μg/l) para diagnosticar este tipo concreto de trombosis en 34 de cada 35 pacientes. Esto le daba una sensibilidad del 97,1%, un valor predictivo negativo del 99,6%, una especificidad del 91,2% y un valor predictivo positivo del 55,7%. Además, el nivel de dímero-D se correlacionaba con la extensión de la trombosis.6 Un estudio posterior, mostraba que el 10% de pacientes con trombosis confirmada tenían un nivel normal de Dímero-D.7
Porque se producen
Las venas del cerebro, ya sean superficiales o profundas, se vacían en los senos venosos durales, que llevan la sangre a la vena yugular y después al corazón. En la trombosis del seno venosos cerebral, los coágulos se forman en las venas del cerebro y los senos venosos. La trombosis de las venas causa infartos venosos, daño al tejido cerebral debido a un congestionado e insuficiente trasporte de sangre. Esto resulta en una edema cerebral, que lleva a pequeñas hemorragias petequiales que pueden convertirse en hematomas mayores. La trombosis de los senos es el mecanismo principal detrás del incremento de la presión intracraneal. Esta condición, sin embargo, no conduce a la hidrocefalia ya que no hay diferencia de presión entre varias partes del cerebro.1 Cualquier coágulo sanguíneo se forma por imbalance entre coagulación y fibrinólisis. Los tres mecanismos mayores para que se produzca esta falta de balance son enumerados en la tríada de Virchow: lesión e Endotelial, lentitud del flujo o estasis sanguínea y los estados de hipercoagulabilidad. La mayor parte de los casos de trombosis cerebrales del seno venoso se produce por hipercoagubilidad.1
Es posible que el coágulo se rompa y migrar(émbolo) a los pulmones, provocando una embolia pulmonar.13 Un análisis de casos previos concluye que este suceso ocurre hasta en un 10% de los casos, y tiene un pronostico muy malo.8
Como se trata.
Varios estudios han investigado el uso de la anticoagulación para suprimir la formación de coágulos sanguíneos en la trombosis del seno venoso cerebral. Antes de que estos estudios se produjesen, existía una preocupación de que áreas pequeñas de hemorragía en el cerebro sangrasen como resultado del tratamiento. El estudio demostró que esta preocupación era infundada.9 Ahora se recomienda la heparina en las primeras fases de tratamiento, seguida de Warfarina, siempre que no haya riesgos de sangrado que hagan este tratamiento inadecuado.31011 Algunos expertos no aprueban el uso de anticoagulantes si la hemorragia es extensiva, en este caso se recomienda repetir el escaneo después de 7-10 días. Si la hemorragia decrece en tamaño, se comienza con los anticoagulantes, mientras que no se les proporciona al paciente si no hay reducción.12
La duración del tratamiento con warfarina depende de las circunstancias y las causas de esta condición. Si la trombosis se desarrolló durante circunstancias temporales (embarazo, por ejemplo) tres meses se consideran suficientes. Si la condición no fue provocada pero no hay causas claras de una forma media de trombofilia, se recomiendan de 6 a 12 semanas. Si hay un desorden severo de trombosis, el tratamiento de trombosis puede ser indefinido.3
La trombolisis (eliminación del coágulo sanguíneo con medicación) se ha llevado a cabo por vía venosa con inyección o directamente en el coágulo por angiografía. Se recomienda que la trombolisis se use solo en pacientes cuya trombosis se deteriores a pesar de una tratamiento adecuado. No está claro qué medicamento y qué modo de administrarlo es el más efectivo. El sangrado cerebral y en otras zonas del cuerpo es una preocupación en el uso de la trombólisis.3 Expertos americanos no recomiendan la trombólisis, pues establecen que se necesita una mayor investigación.3
El incremento de la presión intracraneal, si es severa o amenazante, puede requerir punción lumbar terapéutica, medicación (acetazolamida), o tratamiento neuroquirúrgico. En algunas situaciones, antiepilépticos pueden ser usados para prevenir ataques cerebrales.1
Pronostico.
En 2004, el primero estudio adecuado a gran escala sobre la historia y la prognosis a largo plazo de esta enfermedad fue llevado a cabo. A los 16 meses de recibir diagnóstico, el 57,1% de los pacientes se había recuperado, el 29,5/2,9/2,2% tenían respectivamente síntomas menores/moderados/severos y un 8,3% había fallecido. La muerte era más frecuente en personas con edad sobre 37 años, masculinos, afectados por coma, con desórdenes mentales, hemorragias intracerebrales, trombosis del sistema venoso cerebral profundo, infección del sistema nerviosos central y cáncer.13 Después también se investigó que la mortalidad es del 5,6% durante la hospitalización y del 9,4% en total, mientras el 88% se recuperaron total o casi totalmente.14
En niños con CVST, la mortalidad es del 50%. El resultado se agrava si el niño desarrolla accidentes cerebrovasculares o tiene evidencia de infarto tras un escaneo.15
Epidemiología
La trombosis cerebral del seno venoso es rara, con una estimación de 3-4 incidencias por millón de casos en adultos. A pesar de que puede ocurrir en cualquier grupo de edad, es más común en la tercera edad, donde el 75% de personas que desarrollan CVST son mujeres.3 Ya que estudios antiguos no muestran una diferencia clara de incidencia entre sexos, se cree que el uso anticonceptivos orales en mujeres puede estar detrás de esa disparidad.1 Un estudio de 1995 de Arabia Saudí encontraron una doble incidencia en 7 casos de cada 100.000, algo que se atribuyó al hecho de que el síndrome de Behcet, que aumenta el riesgo de CVST, es más común en Oriente Medio.16Otro estudio de 1973 encontró que la CVST se puede encontrar en autopsias en el 9% de las personas. Muchas de estas eran ancianos con síntomas neurológicos en el período previo a su muerte, y muchos sufrían de insuficiencia cardíaca.17
En niños, un estudio canadiense estableció en 2001 que la CVST ocurre en 6,7 niños por millón anualmente. 43% ocurre en recién nacidos y otro 10% en el primer año de vida. De los recién nacidos, el 84% estaban ya enfermos, principalmente por complicaciones después del nacimiento y la deshidratación.15
En la pandemia de Covid 19 que estamos sufriendo, la vacuna de AstraZeneca ha aparecido varios casos de este tipo de trombo »
El síntoma más frecuente es la cefalea.
Sanidad decidió este lunes suspender de forma «inmediata y preventiva» la vacunación con cualquier lote de AstraZeneca en los próximos 15 días tras detectarse 11 «extraños» casos de trombos cerebrales detectados el fin de semana, uno de ellos en España, y el resto en Alemania (6) y Noruega (4). La directora de la Agencia Española del Medicamento (AEM), María Jesús Lamas, confirmaba que se trataba de un «poco frecuente» trombosis «venosa cerebral» en los «senos venosos», acompañada de una «disminución de plaquetas» y una «activación de una coagulación irregular».
La Sociedad Española de Neurología (SEN) confirma que la trombosis de senos venosos cerebrales es una enfermedad cerebrovascular poco frecuente; de hecho, representa menos del 0,5% de los casos totales de las enfermedades cerebrovasculares que se producen en España. Su incidencia representa en torno a 1 – 1,3 casos por cada 100. 000 habitantes al año.
Aunque puede afectar a cualquier grupo de edad, es ligeramente más frecuente en mujeres y pacientes jóvenes, y entro de los factores de riesgo específicos por sexo están los anticonceptivos orales, el embarazo y el uso de las terapias hormonales.
La SEN especifica que «el síntoma más frecuente de la trombosis de senos venosos cerebrales es la cefalea, presente en nueve de cada diez personas que padecen trombosis de senos venosos cerebrales». La directora de la EMA señalaba ayer que los casos de trombosis de senos venosos cerebrales detectados tras la inoculación de la vacuna contra el Covid de AstraZeneca estuvieron precedidos por un «muy intenso» dolor de cabeza entre 3 y 14 días después del pinchazo que «empeora al tumbarse y que no mejora con analgésicos», además de poder aparecer «vómitos o sangrados irregulares».
Además, la Sociedad Española de Neurología apunta a otros síntomas que pueden ayudar a diferenciar esta cefalea de un dolor de cabeza convencional y que son similares a los de un ictus, como déficits focales, pérdida de fuerza o sensibilidad mantenida, alteración de la marcha o alteración del lenguaje o habla, episodios confusionales o pérdida de visión. Al igual que ha ocurrido con los trombos diagnosticados a personas que recibieron la dosis de AstraZeneca, la trombosis de senos venosos cerebrales también puede provocar vómitos, incluso crisis epilépticas, o rigidez cervical,.
El diagnóstico de este tipo de trombo se realiza mediante pruebas de neuroimagen, como TAC y Resonancia Magnética.
Un dolor de cabeza muy característico
La SEN destaca que el dolor de cabeza que genera una trombosis de senos venosos cerebrales es muy particular. Entre otras características, señala que puede tener un inicio súbito, puede localizarse solo a un lado de la cabeza, empeora cuando el paciente esta recostado o realiza ejercicio, interrumpe el descanso nocturno y, además de no responder a los tratamientos habituales para el dolor de cabeza, empeora de forma progresiva.
A igual que ocurre con todas las enfermedades cerebrovasculares, la trombosis de senos venosos cerebrales se trata de una emergencia neurológica, por lo que cuanto antes se trate al paciente, menores serán sus consecuencias. El tratamiento de elección suelen ser los fármacos antitrombóticos, aunque en casos más graves, la trombectomía endovascular y la trombólisis también son opciones que pueden resultar de mucha utilidad para eliminar el trombo, explica la SEN.
«Generalmente y, sobre todo si se trata a tiempo, los pacientes que padecen trombosis de senos venosos cerebrales tienen un pronóstico favorable. Solo en los casos graves en los que se demore el tratamiento, puede generar discapacidad o tener un desenlace fatal. Además, en los últimos años, gracias también en parte a que la mayoría de los casos de trombosis de senos venosos cerebrales llegan a los hospitales a través del Código Ictus, la rapidez en la atención de estos pacientes es cada vez más rápida y el pronóstico de esta enfermedad cada vez es más favorable», destaca el doctor José Miguel Láinez.
La SEN apunta a que en España la recuperación tras una trombosis de senos venosos cerebrales es completa en aproximadamente un 80% de los pacientes y solo un 5% desarrolla algún tipo de secuela severa. Por lo tanto, el pronóstico de esta enfermedad es mucho más favorable que en otro tipo de enfermedades cerebrovasculares.
Einhäupl K, Bousser MG, de Bruijn SF, et al. (2006). «EFNS guideline on the treatment of cerebral venous and sinus thrombosis». Eur. J. Neurol. 13 (6): 553-9. PMID16796579. doi:10.1111/j.1468-1331.2006.01398.x.
Ferro JM, Canhão P, Bousser MG, Stam J, Barinagarrementeria F (2005). «Cerebral vein and dural sinus thrombosis in elderly patients». Stroke 36 (9): 1927-10. PMID16100024. doi:10.1161/01.STR.0000177894.05495.54.
Dentali F, Gianni M, Crowther MA, Ageno W (2006). «Natural history of cerebral vein thrombosis: a systematic review». Blood 108 (4): 1129-34. PMID16609071. doi:10.1182/blood-2005-12-4795.