La memoria es una función del cerebro que permite al organismo codificar, almacenar y recuperar la información del pasado
La memoria permite retener experiencias pasadas y,
Según el alcance teMPORAL, SE CLASIFICA CONVENCIONALMENTE EN:
MEMORIA A CORTO PLAZO (consecuencia de la simple excitación de la sinapsis para reforzarla o sensibilizarla transitoriamente), MEMORIA A MEDIANO PLAZO Y MEMORIA A LARGO PLAZO (consecuencia de un reforzamiento permanente de la sinapsis gracias a la activación de ciertos genes y a la síntesis de las proteínas correspondientes
El hipocampo es una estructura del cerebro relacionada con la memoria y aprendizaje
En términos prácticos, la memoria (o, mejor, los recuerdos) es la expresión de que ha ocurrido un aprendizaje
Según Carl Sagan, tenemos la capacidad de almacenar en nuestra mente información equivalente a la de 10 billones de páginas de enciclopedia.7 
La memoria está diseminada por distintas localizaciones especializadas. Mientras en algunas regiones del córtex temporal están almacenados los recuerdos de nuestra más tierna infancia, el significado de las palabras se guarda en la región central del hemisferio derecho y los datos de aprendizaje en el córtex parieto-temporal. Los lóbulos frontales se dedican a organizar la percepción y el pensamiento. Muchos de nuestros automatismos están almacenados en el cerebelo.
de memoria como género literario
El psicólogo William James () fue el primero en hacer una distinción formal entre memoria primaria y memoria secundaria (memoria a corto y memoria a largo plazo, respectivamente). Esta distinción reside en el centro del influyente modelo de almacenamiento múltiple de Atkinson y Shiffrin (1968).9
En general, se considera que Hermann Ebbinghaus (1885) fue el pionero en el estudio experimental de la memoria, al haberse utilizado a sí mismo para estudiar fenómenos básicos tales como las curvas de aprendizaje y del olvido e inventar sílabas sin sentido para dicho propósito.
 Memorizar, según :Abenteuer Psyche (Gabriele Amann, Rudolf Wippinger), 1. Auflage, Abbildung 3.16
Memorizar, según :Abenteuer Psyche (Gabriele Amann, Rudolf Wippinger), 1. Auflage, Abbildung 3.16
El almacenamiento Memoria pasa por las siguientes fases:
CODIFICACIÓN O REGISTRO: transformación de la información sensorial en elementos reconocibles y manipulables por la memoria mediante códigos verbales (palabras, números, letras) y códigos visuales (imágenes y figuras). +
ALMACENAMIENTO: Es la retención de la información con el fin de conservarla y recuperarla cuando sea necesario. Según dónde se guarden los datos permanecerán más o menos tiempo en la memoria, desde unos segundos, hasta toda la vida.
RECUPERACIÓN, recordar o recolección: localización y reactualización de la información almacenada. Recordar, es reconocer la información adquirida y almacenada en la memoria. Significa traer a la conciencia la información.10
Registra las sensaciones percibidas a través de los sentidos. Es la fase inicial del desarrollo del proceso de la atención. Esta memoria tiene una gran capacidad para procesar gran cantidad de información a la vez, aunque durante un tiempo muy breve.
Existe una serie de almacenes de información provenientes de los distintos sentidos que prolongan la duración de la estimulación. Esto facilita, generalmente, su procesamiento en la llamada memoria operativa.
Los almacenes más estudiados han sido los de los sentidos de la vista y el oído:
El almacén icónico se encarga de recibir la percepción visual. Es un depósito que almacena, memoria, es una representación isomórfica
Es capaz de mantener aún no se ha reconocido nueve elementos aproximadamente, por un intervalo de tiempo muy corto (alrededor de 250 milisegundos). Los elementos que finalmente se transferirán a la memoria operativa serán aquellos a los que el individuo preste atención, y mantiene almacenados los estímulos auditivos hasta que el receptor haya recibido la suficiente información para poder procesarla definitivamente en la memoria operativa.
Memoria a corto plazo 
El individuo maneja la información a partir de la cual está interactuando con el ambiente.
Es más duradera que la almacenada en las memorias sensoriales, está limitada a aproximadamente 7±2 elementos durante 30 segundos (span de memoria) si no se repasa.
La PRIMICIA Y ULTIMIDAD. Cuando a un grupo de personas se le presenta una lista de elementos (palabras, dibujos, acciones, etc.) para que sean memorizados, al cabo de un breve lapso recuerdan con mayor facilidad aquellos ítems que se presentaron al principio (primicia) o los que se presentaron al final (recencia o ultimidad) de la lista, pero no los intermedios.
El «efecto de primicia» disminuye al aumentar la longitud de la lista, no así el de ultimidad. La explicación que se da a estos datos es que las personas pueden repasar mentalmente los primeros elementos hasta almacenarlos en la memoria a largo plazo, y en cambio no pueden procesar los elementos intermedios. Los últimos ítems, por su parte, permanecen en la memoria operativa tras finalizar la fase de aprendizaje, por lo que estarían accesibles a la hora de recordar la lista. 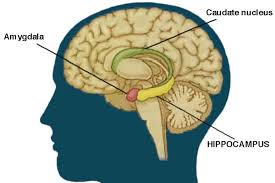
Subsistemas
La memoria operativa o memoria de trabajo está formada por varios subsistemas,:
Sistema Supervisor (el ejecutivo central) y dos almacenes secundarios especializados en información verbal (el lazo articulatorio) y visual o espacial (la agenda visoespacial).
El sistema neuronal ejecutivo central coordina los recursos del sistema y los distribuye por diferentes almacenes, denominados esclavos, según la función que se pretenda llevar a cabo. Se centra, por lo tanto, en tareas activas de control sobre los elementos pasivos del sistema; en este caso, los almacenes de información.
El lazo articulatorio o bucle fonológico, por su parte, se encarga del almacenamiento pasivo y mantenimiento activo de información verbal hablada.
El primer proceso hace que la información se pierda en un breve lapso, mientras que el segundo —repetición— permite refrescar la información temporal.y es responsable de la transformación automática del lenguaje presentado de forma visual a su forma fonológica, por lo que, a efectos prácticos, procesa la totalidad de la información verbal. Esto se demuestra cuando se trata de recordar una lista de letras presentadas de forma visual o auditiva: en ambos casos, una lista de palabras de sonido semejante es más difícil de recordar que una en la que éstas no sean tan parecidas. Asimismo, la capacidad de almacenamiento del «lazo articulatorio» no es constante como se creía (el clásico 7±2), sino que disminuye a medida que las palabras que deben recordarse son más largas.
La agenda visoespacial es el almacén del sistema que trabaja con elementos de carácter visual o espacial. Como el anterior, su tarea consiste en guardar este tipo de información. La capacidad de almacenamiento de elementos en la «agenda visoespacial» se ve afectada —como en el «lazo articulatorio»— por la similitud de sus componentes, siempre y cuando no sea posible traducir los elementos a su código verbal (por ejemplo, porque el «lazo articulatorio» esté ocupado con otra tarea). Así, será más difícil recordar un pincel, un bolígrafo y un lápiz que recordar un libro, una pelota y un lápiz.
Consecuencias de la limitación de recursos
Esta «memoria operativa» afecta la ejecución de varias tareas simultáneas. En las investigaciones de este tipo se dice a un grupo de personas que realicen una tarea principal (por ejemplo, escribir un artículo) y de otra secundaria (por ejemplo, escuchar una canción) al mismo tiempo. Si la tarea principal se realiza peor que cuando se hace en solitario, se puede constatar que ambas tareas comparten recursos.
En líneas generales, el rendimiento en tareas simples empeora cuando éstas requieren la participación de un mismo almacén secundario (por ejemplo, escribir un texto y atender a lo que se dice en la canción), pero no cuando los ejercicios se llevan a cabo de forma separada en los dos almacenes o subsistemas (por ejemplo, escuchar una noticia y ver imágenes por televisión). El procesamiento de información controlado por el «ejecutivo central», la ejecución en ambas tareas se vuelve más lenta, pero no empeora.
Las personas ancianas muestran peor rendimiento en las tareas que requieran el uso del componente del «ejecutivo central» de la memoria de trabajo. Por el contrario, las tareas que precisen del bucle fonológico no se verán tan afectadas por la edad. Memoria a largo plazo
(MLP) es un almacén al que se hace referencia cuando comúnmente hablamos de memoria en general. Es en donde se almacenan los recuerdos vividos, nuestro conocimiento acerca del mundo, imágenes, conceptos, estrategias de actuación, etc.
Dispone de capacidad desconocida y contiene información de distinta naturaleza. Se considera la «base de datos» en la que se inserta la información a través de la «memoria operativa», para usarla posteriormente.
Clasificación por tipo de información]
Una primera distinción considerada dentro de la MLP es la que establece el modelo de Squire en 1992, entre la MEMORIA DECLARATIVA Y LA MEMORIA NO DECLARATIVA conocida como procedimental. La «memoria declarativa» es aquella en la que se almacena información sobre hechos, mientras que la memoria no declarativa nos sirve para almacenar información basados en procedimientos y estrategias que permiten interactuar con el medio ambiente, pero cuya puesta en marcha tiene lugar de manera inconsciente o automática, resultando prácticamente imposible su verbalización.
Memoria no declarativa (implícita)
La «memoria no declarativa» puede considerarse un sistema de ejecución (praxia), implicado en el aprendizaje de distintos tipos de habilidades (se conoce también como memoria procedimental) que no están representadas como información explícita sobre el mundo.11 Este tipo de memoria se activa de manera automática, como una secuencia de pautas de actuación (procedimiento), ante las demandas de una tarea. Consiste en una serie de repertorios motores (escribir) o estrategias cognitivas (hacer un cálculo) que llevamos a cabo de modo inconsciente.
El aprendizaje de estas habilidades se adquiere de modo gradual, principalmente a través de la ejecución y la retroalimentación que se obtenga; sin embargo, también pueden influir las instrucciones (sistema declarativo) o la imitación (mimetismo). El grado de adquisición de estas habilidades depende de la cantidad de tiempo empleado en practicarlas, así como del tipo de entrenamiento que se lleve a cabo. Como predice la «ley de la práctica», en los primeros ensayos la velocidad de ejecución sufre un rápido incremento exponencial, que va enlenteciéndose conforme aumenta el número de ensayos de práctica.
La adquisición de una habilidad lleva consigo que ésta se realice óptimamente sin demandar demasiados recursos de la atención, que pueden usarse en otra tarea al mismo tiempo, de modo que dicha habilidad se lleva a cabo de manera automática.
La unidad que organiza la información almacenada en la «memoria procedimental» es la regla de producción que se establece en términos de condición-acción, y se considera que la condición es una estimulación externa o una representación de ésta en la memoria operativa; la acción se considera una modificación de la información en la memoria operativa o en el ambiente.
Las características de esta memoria son importantes al tratar de desarrollar una serie de reglas que permitan obtener una buena ejecución en una tarea determinada. Esta memoria se explora a través de las praxias, que es la habilidad para poner en marcha programas motores ya aprendidos de manera voluntaria: un movimiento o gesto específico, manejar objetos que requieren una secuencia de gestos, etc.
Memoria declarativa (explícita)
La «memoria declarativa» contiene información referida al conocimiento sobre el mundo y sobre las experiencias vividas por cada persona (memoria episódica). Un ejemplo de la misma es el día de año nuevo, que recordamos y para todos es diferente. También contiene información referida al conocimiento general, sobre todo respecto a los conceptos extrapolados de situaciones vividas (memoria semántica). Mientras que la semántica es más objetiva y un ejemplo de la misma es que color rojo es algo que aprendimos desde pequeños y que para todos es igual, tener en cuenta estas dos subdivisiones de la memoria declarativa es importante para entender de qué modo la información está representada y se recupera diferencialmente.
La «memoria semántica» da cuenta de un almacén de conocimientos acerca de los significados de las palabras y de las relaciones entre estos significados, y constituye una especie de diccionario mental, mientras que la «memoria episódica» representa eventos o sucesos que reflejan detalles de la situación vivida y no solamente el significado.
La organización de los contenidos en la «memoria episódica» está sujeta a parámetros espacio-temporales; esto es, los eventos que se recuerdan representan los momentos y lugares en que se presentaron. Sin embargo, la información representada en la «memoria semántica» sigue una pauta conceptual, de manera que las relaciones entre los conceptos se organizan en función de su significado.
Otra característica que diferencia ambos tipos de representación se refiere a que los eventos almacenados en la «memoria episódica» son aquellos que se han codificado de manera explícita, mientras que la «memoria semántica» posee una capacidad de inferir y es capaz de manejar y generar nueva información que nunca se haya aprendido en forma explícita, pero que se halla implícita en sus contenidos (entender el significado de una frase nueva o de un nuevo concepto utilizando palabras ya conocidas).
También existe la «memoria fotográfica», que es la capacidad de un individuo de recordar hechos incluso pasados muchos años con una precisión increíble. Este tipo de memoria suele ser muy común en personas con síndrome de Asperger.
Memoria inmanente consciente
La refiere a un tipo particular de memoria consistente en la proyección de un subconjunto de memorias de largo plazo sobre la conciencia. La tesis de la memoria inmanente consciente fue postulada por Federico González, psicólogo argentino, quien la definió como:
Una proyección de la memoria de largo plazo episódica sobre la conciencia, realizada momento a momento. La tesis de la memoria inmanente (MIC) consciente sostiene que un fragmento compactado de información proveniente del sistema episódico de memoria de largo plazo (MLP) se encuentra proyectado sobre la conciencia, a modo de una macro huella mnémica. La MIC es experimentada conscientemente como un background que confiere significado a la experiencia actual. En tal sentido, la conciencia resulta una síntesis que amalgama simultáneamente la memoria con la experiencia presente. En consecuencia, desde la perspectiva de la MIC, la conciencia constituye la condensación integradora de un subconjunto de la memoria de largo plazo unido a la conciencia perceptual e ideativa presente. En síntesis, la MIC es una subestructura de la mente que comprende un conjunto de experiencias pasadas que se presenta reflejado o «incrustado» sobre la conciencia.1213
La tesis de la memoria inmanente consciente ha sido aplicada para explicar un vasto conjunto de fenómenos de la vida mental tales: algunos aspectos del tiempo subjetivo como la sensación de duración y la dimensionalidad de antigüedad de los recuerdos, la noción de identidad personal, la naturaleza de las retenciones y protenciones fenomenológicas y la orientación generalizada de la realidad espacio-temporal.
La tesis de la memoria inmanente consciente asume que la conciencia presente contiene mucha más información que la que se le atribuye en el almacén de corto plazo (ACP). También relativiza el postulado que sostiene que para fijar nueva información en la memoria de largo plazo es necesario realizar un repaso mental interno de la misma. Por el contrario, se sostiene la hipótesis de un mecanismo de autoguardado permanente que permitiría almacenar síntesis de experiencias episódicas sin que se busque deliberadamente hacerlo.
La Memoria inmanente consciente también se ha aplicado para comprender algún tipo de sueño donde en la vivencia del soñante se dan por supuestos hechos que, en rigor, no ocurrieron durante el sueño.
Los recuerdos
Son imágenes del pasado que se archivan en la memoria. Nos sirven para traer al presente algo o a alguien. Se definen también como una reproducción de algo anteriormente aprendido o vivido, por lo que están vinculados directamente con la experiencia.
Las conexiones neuronales de la corteza son estimuladas por la información y se fortalecen y vinculan al contexto emocional en que se formaron. Una vez codificadas reside en la región de la corteza en la que la información fue percibida y procesada. Cuando sea necesario, o suscitado por la emoción, esa memoria se activa para utilizarse como memoria de trabajo.14
Según el psicoanálisis, el aferrarse a un recuerdo puede generar depresiones y, en casos extremos, incluso una ruptura con la realidad actual.
Los recuerdos de un colectivo humano nos dan una aproximación más cercana de la realidad que la propia historia, puesto que ella suele saltarse los hechos individuales para centrarse en los acontecimientos globales.
Desde la fenomenología, el filósofo Paul Ricoeur también explica que el recuerdo es una imagen. Al recordar, como suele decirse, representamos un acontecimiento pasado,15 constituyendo así una estructura común entre memoria e imaginación en el momento que tiene lugar en nuestra mente la representación de algo ausente.
Patologías
La enfermedad del Alzheimer es una clase de demencia progresiva causada por la aparición de placas y nudos neurofibriliares en distintas regiones de la corteza cerebral y el hipocampo. Al principio, estos nudos y placas, se concentran en la región temporal media, esta zona está relacionada con el establecimiento de nuevas memorias explícitas. Esta zona es vital para el establecimiento de la memoria episódica y contribuye a la formación de nuevas memorias semánticas.
La apolipoproteína E (APOE) es esencial para el catabolismo de lipoproteínas. Además, ha sido vinculada a enfermedades cardiovasculares. La APOE E4 ha sido relacionada con una mayor sensibilidad a desarrollar Alzheimer. El APOE4 tiende a producir una acumulación amiloide en el cerebro, algún tiempo antes de que aparezcan los primeros síntomas del Alzheimer. A pesar de los recientes hallazgos, la presencia del gen apolipoproteína E, no permite explicar todos los casos de la enfermedad del Alzheimer.
Alteraciones cuantitativas
Amnesia
La mala memoria está provocada por factores o deficiencias biológicas ajenas propiamente al funcionamiento normal de la misma.16La amnesia es la ausencia de recuerdos de un período determinado de la vida. El sujeto suele estar consciente de que son recuerdos que existieron, pero que se han perdido. Pueden ser parciales o totales.
Amnesia parcial: afectan los recuerdos de un campo reducido de memoria visual, auditiva o verbal. Puede estar presente en trastornos orgánicos del cerebro, lesiones de la corteza cerebral por traumatismos, deficiencia circulatoria, intoxicaciones o trastornos psicogénicos. Si la amnesia es de etiología orgánica, suele ser definitiva, mientras que la amnesia temporal de etiología psicogénica suele ser transitoria.
Amnesia total: es la que se vuelve extensiva a todos los elementos y formas de conocimiento, que corresponde a un lapso determinado de la vida del sujeto. Según la cronología del lapso olvidado, se divide en:
Anterógrada o de fijación: incapacidad de evocar hechos recientes, pero si logra recuerdos antiguos. Suelen ser transitorias, pero pueden convertirse en definitivas, como ocurre en las demencias.
Amnesia retrógrada o de evocación: es la dificultad para evocar el recuerdo de vivencias conservadas del pasado y que en otras oportunidades han podido recuperarse.
Amnesia global o retroanterógrada: afecta simultáneamente la fijación de eventos presentes y la evocación de recuerdos pasados. Se observa en los períodos terminales de las demencias.
Hipomnesia
Es la disminución de la capacidad de la memoria debido a una dificultad tanto en la fijación como en la evocación. Se observa en personas psiquiátricamente sanas con preocupaciones profundas que acaparan la atención, así como en pacientes con neurosis.
Hipermnesia
Es el aumento o hiperactividad de la memoria, frecuente en pacientes maníacos o delirantes, y se presenta también en sujetos con entrenamiento especial de la memoria.
Dismnesia
Es una alteración cuantitativa que traduce siempre en una disminución de la memoria, imposibilita evocar un recuerdo en un momento dado y evoca otros en forma borrosa o poco nítida.
Se llama así por la dificultad para evocar un recuerdo en determinado momento, pero que más tarde puede ser evocado espontáneamente. Esto ocurre en las personas normales en forma esporádica, al tratar de recordar nombres propios, fórmulas, etcétera. Este es uno de los síntomas iniciales que se da en la senectud. Y en forma permanente en el comienzo de la demencia.
Alteraciones cualitativas
Se han agrupado bajo la denominación de paramnesia, es decir, los falsos reconocimientos o recuerdos inexactos que no se ajustan a la realidad. Los principales son:
Fenómeno de lo ya visto (déja vu): es la impresión de que una vivencia actual ha sido experimentada en el pasado y en la misma forma. Se puede observar en personas sin ningún padecimiento mental o en sujetos con neurosis o con esquizofrenia.
Fenómeno de lo nunca visto (jamais vu): sensación de no haber visto o experimentado nunca algo que en la realidad ya se conoce.
Ilusión de la memoria: es la evocación deforme de una vivencia, al cual se le agregaron detalles creados por la fantasía. Se observa en personas sin padecimientos mentales y en sujetos con delirios o con esquizofrenia.
Tratamientos contra la pérdida de la memoria
Algunas experiencias e investigaciones sugieren que el consumo de Ácido graso omega 3 tiene efectos benéficos sobre el desarrollo del cerebro, y en procesos tales como la memoria y la concentración.17 También hay estudios que sugieren que el consumo de omega 3 durante el embarazo puede tener una buena influencia en el bebé18 e incluso grupos de niños en edad escolar aumentaron notablemente su rendimiento después de ingerir pastillas con aceite de pescado rico en omega 3.
Otra de las ayudas eficaces para mejorar la retención consiste en dedicar una buena parte del tiempo de estudio no a la lectura sino al repaso mental de lo que se ha leído, a su reproducción ordenada sin recurrir al libro más que cuando falla la memoria. De esta forma no solo la retención, sino los mecanismos de captura de lo retenido, se consolidan y perfeccionan. En algún caso, el tiempo dedicado con fruto a este repaso mental ha llegado hasta el 80 %.16
Se ha demostrado la asociación entre los trastornos de la memoria con el consumo de gluten, tanto en personas con enfermedad celíaca como con sensibilidad al gluten no celíaca (pruebas negativas para enfermedad celíaca pero mejoría al eliminar el gluten de la dieta). Si bien su papel es controvertido, la dieta sin gluten parece ejercer un efecto protector sobre las alteraciones de la memoria, más efectivo cuanto menor sea el retraso desde el comienzo de los primeros síntomas.19 (Véase también Trastornos neurológicos relacionados con el gluten).
Mecanismos de olvido
Olvidar es algo normal, incluso necesario, pues nos evita acumular un exceso de datos inútiles. Imagine por un momento que fuese capaz de recordar absolutamente todo lo que ha aprendido y vivido a lo largo de su vida. Por lo tanto, se trata de acordarse únicamente de lo importante.
El olvido puede deberse a varias causas:
Caducidad: Los datos almacenados pueden ir diluyéndose con el paso del tiempo. Y esto tiene un sentido en la memoria sensorial, y en los plazos corto y mediano, pues es la manera de que no lleguen a saturarse.
No encontramos explicación al hecho de que se olviden cosas que estaban almacenadas en la memoria a largo plazo, pues su capacidad es prácticamente ilimitada. Algunos investigadores afirman[cita requerida] que los recuerdos se conservan de por vida, y lo que falla es el modo de acceder a ellos.
Es conocido el efecto agenda desde la salida de las agendas electrónicas, hoy reforzado por el Efecto Google que es la tendencia a olvidar información que se puede encontrar en Internet utilizando motores de búsqueda, en lugar de esforzarse en recordarla.20
Este efecto de olvido producido al navegar en la red también aparece al utilizar otras tecnologías como las cámaras de fotos. En un experimento llevado a cabo en el Museo de Arte de la Universidad de Fairfield, la Doctora en Psicología Lady A. Henkel descubrió que los visitantes que tomaban fotos de las obras expuestas tenían un peor recuerdo tanto sobre qué objetos habían visto como de los detalles de estos, en comparación con aquellos visitantes que se habían limitado a observar. Esto ocurrió a pesar de que ambos grupos de visitantes dedicaron el mismo tiempo a visualizar cada obra. La explicación a estos resultados es que, en muchas ocasiones, las personas confiamos en la tecnología (una cámara de fotos en este caso) para que almacene parte de los recuerdos por nosotros. Este efecto de olvido se podría contrarrestar si, más adelante, dedicásemos un tiempo suficiente a observar de nuevo las fotografías tomadas, lo cual reforzaría el recuerdo de aquellos momentos, fortaleciéndolos en la memoria a largo plazo.21
Problemas de acceso: A veces no podemos acceder al contenido de nuestra memoria, especialmente si el estrés nos hace producir hormonas (glucocorticoides) que bloquean la función de acceso. Mediante algunos ejercicios de memoria podemos aumentar la posibilidad de que esto no suceda.
Eliminación: Aparece en el caso de informaciones dolorosas, frustrantes y molestas y cuando se han vivido situaciones extremas o traumáticas. Frente a este mecanismo de olvido los ejercicios de memoria no sirven para nada. De todos modos, algunas experiencias y recuerdos desagradables pueden emplearse para mejorar la memoria.
Los 7 pecados de la memoria (según Schacter)[
1. Paso del tiempo
2. Distracción: Despistes que, según el autor, son responsabilidad de más bien la falta de atención.
3. Bloqueo: ‘Lo tengo en la punta de la lengua’.
Por comisión:
4. Atribución errónea: Atribuir un recuerdo a una fuente errónea. Por ejemplo, atribuirnos ideas que en realidad no son nuestras.
5. Sugestionabilidad: Los recuerdos se ven influidos por agentes externos. Por ejemplo, no nos acordamos bien de lo que sucedió un día cualquiera. Pero a medida que un amigo te lo cuenta, lo recuerdas como tal, aunque haya datos falsos.
6. Sesgo: El recuerdo se ve influido por nuestro estado (sentimientos, punto de vista) actual.
7. Persistencia: Permanencia de recuerdos que desearíamos olvidar.22
La memoria y la psicología
La memoria es una función cerebral que interviene en todos los procesos de aprendizaje del ser humano. Es vital para la supervivencia del individuo como lo ha sido para la supervivencia de la especie. Esto es un punto común con muchas de las especies animales, por no decir todas y cuando consideramos novedosos aspectos en estudio, como la memoria de las células, también es un punto común con las plantas y los demás seres vivos.
La memoria humana, al igual que el ser humano en sí, es compleja y fascinante. Es a ella a la que mayor atención préstamos y a la que mayor esfuerzo le exigimos en nuestras vidas ya que, nuestra vida existe gracias a nuestra memoria. La vida está formada por recuerdos. La memoria es la capacidad de adquirir, almacenar y recuperar información. Somos quienes somos gracias a lo que aprendemos y recordamos. Sin memoria seríamos incapaces de percibir, aprender o pensar, no podríamos expresar nuestras ideas ni tendríamos una identidad personal, porque sin recuerdos sería imposible saber quiénes somos y nuestra vida perdería sentido. Este mapa conceptual consiste en un resumen del tema de la memoria humana que estamos estudiando. Del título principal «La memoria humana» salen distintos apartados en los que se divide el tema.
En primer lugar se encuentra la «complejidad de la memoria» en el que se explica en qué consiste la memoria y las investigaciones que se han llevado a cabo sobre ella tanto las que estudian la repetición, los esquemas y la memoria a corto plazo. Otro de los apartados es el de la neuropsicología de la memoria que ya habla sobre investigaciones más recientes y científicas sobre nuestra memoria y los procesos que tienen lugar en nuestro cerebro para que esta funcione correctamente.
Los dos últimos apartados que aparecen son los de los procesos básicos de la memoria, en el que se nombran las tres funciones básicas de ésta, y la estructura y funcionamiento de la memoria, en el que se nombran los tres sistemas de memoria que se comunican e interactúan entre sí. Estos sistemas fueron reconocidos por los psicólogos Richard Atkinson y Richard Shiffrin mediante el desarrollo de la teoría multialmacén de la memoria.
Hay fallas en el proceso y la función de la memoria que no son propiamente olvidos o dificultades en el almacenamiento o la recuperación, sino que son distorsiones de la información. Algunos de estos trastornos alcanzan un grado de enfermedad ya y son llamados «paramnesias»; otros por el contrario son bastante poco frecuentes o leves en su ocurrencia. En ambos casos las manifestaciones pueden ser similares, pero diferir en la intensidad o frecuencia.
Entre las anomalías más frecuentes encontramos:
«Tu cara me parece conocida».
“Olvidar el nombre”
“Sensación de conocer”
«Lo tengo en la punta de la lengua
Laguna temporal: es cuando se olvidan algunos fragmentos de algo o lo ocurrido en un lapso específico, por lo general cuando en ese lapso no ocurre nada relevante y se estaban ejecutando funciones o labores sobre aprendidas. Ejemplo, cuando olvidamos parte del recorrido habitual a nuestra casa: no recordamos cuando pasamos por determinado punto (por el que tuvimos que haber pasado).
Falsificación de la memoria o falsos recuerdos: el aparato psíquico crea recuerdos para llenar lagunas en la memoria. Este tipo de trastorno tiende a ser altamente problemático para el sujeto y merecer especial cuidado desde la psicología.
Deja vu: es una anomalía del reconocimiento que implica que experimentamos esa situación de «esto ya lo he visto» o «esto ya lo he vivido», aún a sabiendas de que es la primera vez que lo vemos o vivimos.
Jamais vu: es el caso contrario al anterior. Aquí, aunque el individuo conoce y sabe que conoce determinada situación y la recuerda, no experimenta ninguna sensación de familiaridad.
Referencias
↑ Feldman, Robert S. (2005). Psicología con aplicaciones en países de habla hispana. México: McGraw Hill. ISBN 978-607-15-0287-2.
↑ Esteban, Ana Isabel (24 de julio de 2018). «Solidaridad Intergeneracional». MEMORIA, FUENTE DE VIDA. Consultado el 18 de enero de 2021.
↑ Córdoba-Montoya, D. A., J. Albert, S. López-Martín. «Potenciación a largo plazo en la corteza humana». www.neurologia.com. Consultado el 8 de diciembre de 2017.
↑ Kandel, E. (1997). Neurociencia y conducta. Pearson. ISBN 978-84-89660-05-2.
↑ Tres14 (2009). Memoria (Programa de televisión). Archivado desde el original el 21 de septiembre de 2010. «Entrevista a Ignacio Morgado, catedrático de psicobiología en la Universidad Autónoma de Barcelona ».
↑ www.saludparati.com. «Cómo Mejorar tu Memoria». Archivado desde el original el 3 de enero de 2010. Consultado el 22 de diciembre de 2009.
↑ Morgado Bernal, Ignacio (2005). «Psicobiología del aprendizaje y la memoria». Cuadernos de Información y Comunicación 10. ISSN 1135-7991. Consultado el 19 de diciembre de 2009.
↑ García-Allen, Jonathan (2016). «Tipos de memoria: ¿cómo almacena los recuerdos el cerebro humano?». Psicologiaymente.net. Consultado el 5 de septiembre de 2016.
↑ Granell, Abel (2011). «Bloque IV. Procesos psicológicos II: la memoria». Apuntes de Psicología.
↑ Carboni Román, Alejandra (2007). «Desarrollo de la memoria declarativa». EduPsykhé: Revista de psicología y psicopedagogía 6 (2): 245-268. ISSN 1579-0207.
↑ Federico González (1912). Editorial académica española, ed. «Ensayos sobre el tiempo subjetivo».
↑ González, Federico. «Memoria inmanente consciente». YouTube/Psych Life.
↑ National Geographic. Features. Mappig Memory in 3D. Consultado el 5 de junio de 2012.
↑ Aranzueque, Gabriel (1997). «Paul Ricoeur: Memoria, olvido y melancolía». Revista de Occidente (198): 105-122. ISSN 0034-8635.
↑ Saltar a:a b Pinillos, José Luis (1977). Introducción a la psicología. Madrid, España: Alianza Universal. ISBN 84-206-2100-5.
↑ Innis, Sheila M. (2008). «Dietary omega 3 fatty acids and the developing brain». Brain Research (en inglés) 1237: 35-43. doi:10.1016/j.brainres.2008.08.078.
↑ Palmer, D. J.; Sullivan, T.; Gold, M. S.; Prescott, S. L.; Heddle, R.; Gibson, R. A.; Makrides, M. (2012). «Effect of n-3 long chain polyunsaturated fatty acid supplementation in pregnancy on infants’ allergies in first year of life: randomised controlled trial». BMJ (en inglés) 344. doi:10.1136/bmj.e184.
↑ Makhlouf, S., Messelmani, M., Zaouali, J., Mrissa, R. (15 de diciembre de 2017). «Cognitive impairment in celiac disease and non-celiac gluten sensitivity: review of literature on the main cognitive impairments, the imaging and the effect of gluten free diet». Acta Neurol Belg (Revisión). PMID 29247390. doi:10.1007/s13760-017-0870-z.
↑ «El ‘efecto Google’ afecta al uso de la memoria». El País. 15 de julio de 2011.
↑ Henkel, Linda A. (5 de diciembre de 2013). «Point-and-Shoot Memories: The Influence of Taking Photos on Memory for a Museum Tour». Psychological Science (en inglés) 25 (2): 396-402. doi:10.1177/0956797613504438.
↑ Schacter, Daniel (2001). Los siete pecados de la memoria. Houghton Mifflin Hancourt. ISBN 978-061-80-4019-3.










 40 años de sida: el primer caso de VIH del Estado se diagnosticó en Vall d’Hebron
40 años de sida: el primer caso de VIH del Estado se diagnosticó en Vall d’Hebron