ChatGPT:

QUE ES ChatGPT
Hace como 20 años, esto hubiera parecido de Julio Verne, ahora es verdad, le pides a una maquina, que opine sobre el vestido que te pondrás en una fiesta y te lo dice con su lógica, con su experiencia y su cultura.
Vamos esto es “La de Dios”
Se trata de uno de los sistemas de IA INTELIGENCIA ARTIFICIAL, capaz de responder a cualquier cosa que le solicites.
Se trata de una inteligencia artificial que está entrenada para mantener conversaciones.
EL ChatGPT es un sistema de chat basado en el modelo de lenguaje por Inteligencia Artificial GPT-3, desarrollado por la empresa OpenAI. Es un modelo con más de 175 millones de parámetros, y entrenado con grandes cantidades de texto para realizar tareas relacionadas con el lenguaje, desde la traducción hasta la generación de texto.
A una inteligencia artificial se la entrena a base de texto, se le hacen preguntas y se le añade información, de manera que este sistema, a base de correcciones a lo largo del tiempo, va «entrenándose» para realizar de forma automática la tarea para la que ha sido diseñada. Este es el método para entrenar a todas las IA, tanto a la de ChatGPT como otras del estilo de los Magic Avatars de Lensa.

EL ChatGPT, esta IA ha sido entrenada para mantener conversaciones con cualquier persona. Sus algoritmos deberían ser capaces de entender lo que le estés preguntando con precisión, y de responderte de una manera coherente.
Es capaz de dar respuestas acertadas y completas, incluso de varios párrafos. Es capaz de expresarse de manera natural y con información muy exacta, lo que hace muy complicado distinguir que el texto ha sido generado por IA.
Puedes pedirle una redacción de 1000 palabras sobre un tema concreto, y la IA te la generará antes de que tú hayas tenido tiempo de abrir Google para buscar el primer concepto. Sin embargo, como cualquier modelo de IA, es posible que cometa errores .
Esta IA podría acabar con Google y buscadores similares. Sin embargo, en muchos temas es poco precisa, sobre todo en nombres y algunos conceptos, por lo que todavía no está a la altura de permitirte copiar lo que ha escrito y pegarlo.
Además de responder a la pregunta, esta AI tiene un sentido del contexto y reconoce todo lo que habéis estado hablando hasta ahora, por lo que si le haces alguna pregunta relacionada con una respuesta que te ha dado, sabrá identificar si te refieres a ello sin tener que darle toda la explicación.

Hemos entrevistado a GPT-3 sobre cómo será la vida en 2030 y esto es lo que nos ha dicho de Marte y el futuro (y dice que se llama Luis)
Cómo usar ChatGPT
Empezar a tener tus conversaciones con esta inteligencia artificial es muy sencillo. Primero entrar a su web oficial, que es https://chat.openai.com.
La primera vez que entres tendrás que crearte una cuenta en la web de OpenAI, pero todo es completamente gratuito, y también el chat.
Y una vez inicies sesión ya entrarás en el chat. En él, abajo del todo tendrás la barra donde tú escribes lo que quieras. Antes de empezar verás algunos ejemplos en inglés de cosas que le puedes preguntar, pero recuerda que también puedes hacerle preguntas en Español, y pedirle que te hable en español o en cualquier otro idioma que prefieras.
Es fácil usar ChatGPT en español.
El ingenio a la hora de realizar preguntas o peticiones puede ser clave para obtener resultados sorprendentes.
Todo lo que escribas quedará registrado, y podrá ser revisado después por los desarrolladores de OpenIA para seguir entrenando a ChatGPT.
Es conveniente no incluir información personal ni peticiones que sean potencialmente peligrosas o delictivas.
Qué puedes hacer con ChatGPT

ChatGPT es una de esas herramientas cuyas funciones completas todavía no han sido descubiertas, porque en parte depende del ingenio de las personas que interactúan con ella.
Lo más sencillo es pedirle que te explique cualquier cosa, evento o concepto, y la IA lo hará a través de los datos con los que se la ha entrenado.
Esto también te servirá para que te escriba artículos o resumenes pudiendo pedirle un número máximo de caracteres o palabras explicando sucesos históricos, personajes famosos, dispositivos tecnológicos.
Se comporta como periodista particular, o una chuleta para clases.
También puedes pedirle que te escriba estos textos de una manera determinada. Por ejemplo, puedes pedirle que te haga un guión de YouTube o de TikTok para explicar estas cosas, que lo haga con un tono concreto como informal o más serio, o incluso que lo haga con las tonalidades de ciertas regiones. Escríbelo como un andaluz.

EN GENBETA
Esta IA ha llegado para acabar con Google, StackOverflow y hasta con los tutoriales web: esto es lo que puedes preguntar a ChatGPT
También puedes pedirle líneas de código, fichas de especificaciones de productos, comparativas, etcétera. También puedes pedirle que te escriba poemas, chistes o letras de canciones. Puedes pedir que realice textos como si le hablara a un niño de 5 años, y en definitiva, cualquier cosa relacionada con el lenguaje o que pueda ser expresada a través de él.
También puedes pedirle muchas otras cosas, como listas de páginas web o de herramientas, pedirle consejo sobre qué móvil comprar, o temas relacionados con la cultura general, trivials, traducciones, definiciones, explicaciones, y todo lo que se te ocurra. Hay muchos ejemplos, y puedes intentar preguntarle cosas que nadie haya preguntado antes para descubrir nuevas funciones.
La integración de ChatGPT en Bing (el buscador de Microsoft) no es una sorpresa. La empresa con sede en Redmond ya ha expresado en varias ocasiones su interés por incorporar el software de generación de imágenes de OpenAI, DALL-E 2.
No es la única vez que ha mostrado su interés por la empresa de IA. En 2019, Microsoft respaldó a la compañía fundada por Elon Musk y Sam Altman, entre otros, ofreciendo mil millones de dólares en fondos. De hecho, ambas tecnológicas habían formado una asociación de varios años para desarrollar tecnologías de supercomputación de inteligencia artificial en el servicio de computación en la nube Azure de Microsoft, recuerda Bloomberg.

Representación de ChatGPT como un androide hecha por la IA Dall-E de OpenAI
REDACCIÓN / Terceros
ChatGPT ha revolucionado el mercado técnológico. Los especialistas aseguran que el chatbot de inteligencia artificial de OpenAI presenta unas prestaciones únicas, sobre todo por su capacidad de conversar con humanos. Trabaja con 175 millones de parámetros y está entrenado a conciencia con grandes cantidades de texto para completar todo tipo de tareas relacionadas con el lenguaje, desde la generación de cualquier redacción hasta la traducción. De hecho, La Vanguardia mantuvo una entrevista con este bot.
Su eficacia lo convierte en un competidor potencial de Google. La compañía con sede en California no quiere quedarse atrás en esta carrera y estaría trabajando en desarrollar una tecnología similar en su sistema Language Model for Dialogue Applications (LaMD
APRENDIZAJE AUTOMÁTICO
La llegada de un modelo de Inteligencia Artificial que redacta como un humano y resuelve problemas está impactando ya en los colegios y las universidades
Hace todos las tareas que le pides», aseguran los estudiantes
Las universidades ven futuras ventajas, pero no saben cómo van a abordar el problema del plagio a partir de ahora

Ambiente de estudiantes universitarios en la biblioteca del edificio histórico de la UB en plaza Universidad
Ana Jiménez
FRANCESC BRACERO, CARINA FARRERAS
BARCELONA, BARCELONA
Lo preparo en un minuto”, respondió Aran a su profesora.
La docente, Ester Robles, había expresado su preocupación por el artículo mensual de la revista del instituto que la clase debía entregar coincidiendo con la semana de exámenes. Aran propuso utilizar una nueva herramienta de Inteligencia Artificial (IA), ChatGPT, capaz de elaborar textos de forma automática. La profesora sintió curiosidad por lo que le relataba el alumno con tanta pasión, aunque mantenía cierto escepticismo sobre el resultado.
-“¿Un artículo resumen con opinión en 150/250 palabras va bien?”
El artículo debía recoger la charla que el fundador de Open Arms, Óscar Camps, había impartido en el instituto (el Vicens Vives de Girona). Aran Oliveras le puso un enunciado con palabras claves y una breve valoración y ¡voilà!, en menos de un minuto apareció un artículo. “Estaba bien redactado, contenía las ideas principales, bien ordenadas, con una valoración adecuada. Era casi indistinguible respecto a lo que hubiera redactado un buen alumno”, admite Robles.
“Los profesores estamos fascinados con esta nueva IA y preocupados al mismo tiempo, porque nos damos cuenta de las posibilidades que se nos abren pero también de los riesgos que entraña”. Aún no se ha hablado en el claustro, pero el conocimiento de esta herramienta por parte de la profesora de lengua castellana sí ha tenido consecuencias: de momento, los textos descriptivos (que entran en selectividad) se harán en clase y no en casa.
ChatGPT es el tema estrella en las salas de profesores de los centros educativos. Se trata de una herramienta de IA “superdotada”, cuyo uso se abrió al público de forma gratuita y viralizó el pasado diciembre. Como puede movilizar conocimientos depositados en internet –de momento, sólo hasta 2021–, tiene una respuesta tan rápida, eficiente, ordenada, correcta de cualquier disciplina, que ha puesto a temblar no solo las paredes de colegios sino también las de las universidades.
El principal temor es el riesgo de plagio. Su irrupción, que era previsible pero no a corto plazo, ha pillado a todos desprevenidos.
No es El rincón del vago ni una búsqueda de Google. El chatbot responde a cualquier cuestión que se le plantee, incluso a la más disparatada, y se le puede pedir que profundice sobre la idea si no se obtiene el resultado previsto. Ciertamente, ¿cómo saber si lo escrito corresponde a un trabajo y pensamiento propio o es el resultado de consultar al bot?
Su uso no parece estar extendido aún entre los estudiantes, pero ya hay ejemplos de que se utiliza. En la Pompeu Fabra ya les consta un trabajo de clase realizado por IA. Fue aprobado con buena nota. El profesor tuvo conocimiento del plagio a través de un anónimo y el campus está dirimiendo cómo actuar al respecto, al margen de la sanción prevista para estos casos. El uso de la IA está presente en otras universidades, según ha comprobado La Vanguardia, aunque, entre las consultadas, no tengan conocimientos de incidentes.
“Mejora este ensayo filosófico”, pide una estudiante de grado de Filosofía de la UB. Y el bot le devuelve incorrecciones y muchas sugerencias de mejora. “Resuelve esta práctica en el lenguaje Python”, propone un estudiante de Matemáticas del mismo campus.
“Hay que revisar los resultados porque no siempre los da de forma correcta, pero le pasas la computadora para comprobarlo y le vuelves a dar, y el chat se disculpa por haberlo hecho de forma incorrecta y te lo da correcto”, apunta el futuro matemático que se enteró por el grupo de WhastApp de la clase.
“Es muy loco porque es muy bueno, estamos alucinando”, afirma una compañera de clase a quien el bot empezó a contestarle en castellano, pese a los enunciados en catalán, y ahora, un mes después, ya lo hace en catalán.
¿Cuáles son las fases de una cirugía de pectum excavatum ?”, le pregunta Xavi, un estudiante de Medicina de la UAB tras asistir a una práctica de quirófano de una operación de esternón en el hospital. “Yo lo haría igual, pero su información me ayuda a empezar y ya solo tengo que añadir algunos comentarios”.
Esta revolución no ha hecho más que empezar. “Aún está en versión beta y tiene información hasta 2021, por tanto mejorará, pero ya puede elaborar un trabajo de fin de grado o de fin de master con pocos fallos”, asegura el ingeniero informático Senén Melquíades Palanca. Y es que puede hacer muchas cosas. “Plantea los índices, resume información, responde dudas, mejora textos y sugiere cómo puedes hacer la presentación verbal…»
“¿Qué son los patrones de diseño? ¿cómo se implementan y cuáles debo utilizar para mi caso concreto?», le preguntó Senén. «Son operaciones informáticas complejas que hubiera tardado tiempo en lograr un resultado. Me dio el código en minutos”, afirma.
¿El bot responde con suficiente calidad? A juicio de los estudiantes, algunas respuestas son excelentes, otras mejorables y otras contienen errores por lo que conviene revisarlo siempre. Depende también del área de conocimiento. “En una prueba de la facultad contestó bien 3 diagnósticos de los 9 que nos presentaron”, indica el alumno de Medicina. «Yo lo uso como asistente en mi aprendizaje, más que para hacer trabajos que prefiero hacer yo porque luego tengo que ir a examen».
Este diario probó el Bot con algún ejercicio de las pruebas de acceso a la universidad de 2022 en Catalunya (ejercicios de física, matemáticas y filosofía) y el resultado fue desde excelente a solo correcto, comparándolo con las correcciones que el propio Departamento de Universidades cuelga en su web.
Las universidades parten de un planteamiento positivo: “esto ha venido para quedarse y permitirá mejorar la forma de enseñar y de aprender”. Supone, indican, un salto cualitativo pedagógico que los profesores deben dar en poco tiempo, un salto equiparable por su dimensión a la adaptación tecnológica de los trabajadores durante la pandemia.
Consideran que se superará, como cuando apareció un buscador como Google, correctores ortográficos, diccionarios en línea o calculadoras científicas. Ahora ya nadie cuestiona estos avances. A cambio, habrá beneficios, por ejemplo en los materiales que ahora los profesores no pueden presentar por temas de copyright, y que podrán pedir a la IA que genere originales (fotografías, dibujos, esquemas).
La educación lo apropiará en su beneficio. Pero la transformación pedagógica que supone será un reto para el profesorado. «En el aula, mientras habla el profesor, yo le voy preguntando a la IA dudas que me surgen y si no las entiendo le pido que lo explique de forma más sencilla», apunta la estudiante de la UB.
Por el momento, la inquietud perentoria de las instituciones es la evaluación. Tanto en colegios como en universidades. Los chicos de 1º de bachillerato están empezando este mes el trabajo de investigación que tienen que presentar en 2º de bachillerato. Un trabajo que requiere un esfuerzo para encontrar el planteamiento, la búsqueda de información, la elaboración y las conclusiones. Todo esto se le puede pedir al chat Bot, incluso por fases, incluso adecuado a la edad, incluso con algunos errores. «Esto nos lo cambia todo, y nos lo cambia ya, no el próximo curso, ¿cómo planteamos ahora estos trabajos?”, se pregunta la profesora Isabel López, del instituto de Salesianos de Terrassa.
Los colegios cuentan con la ventaja de la presencialidad y que los profesores conocen a sus estudiantes. En las universidades esto es más difícil (aún más en las online).
Por una parte, los trabajos que hacen fueran del aula son un elemento clave en la evaluación continua que promovió el Plan Bolonia. Por otra, en la entrega de sus titulaciones se juegan su prestigio. Por ello invierten ingentes cantidades de dinero en programas avanzados antiplagio (que no cubren la nueva IA porque los textos son originales).
Los trabajos individuales o en grupo son un instrumento clave de la evaluación continua. Sustituirlos por exámenes en clase requiere cambios organizativos (y ya constan en las programaciones de cada grado o master). Y comprobar que el alumno ha adquirido conocimiento con pruebas orales sobre los trabajos implica mayor tiempo destinado a esa tarea y, por tanto, aumento de coste económico para los campus.
Ante la sombra de la amenaza, algunos centros académicos están reaccionado de forma defensiva. Según The Guardian , las ocho principales universidades australianas han revisado la metodología de las evaluaciones en 2023. Básicamente, más exámenes “a papel y bolígrafo”.
En Nueva York han prohibido su uso en las escuelas. En Catalunya, el Departamento de Educación no va prohibirlo “en todo el sistema y para todos, ya que esto sería una medida poco efectiva”. Según fuentes de la conselleria, es mejor plantear a los centros que eduquen en el uso de la IA, “que puede aportar muchísimo conocimiento y ventajas”.
En todo caso, recuerdan que las escuelas tienen, por ley, autonomía organizativa, de gestión y pedagógica, por tanto, cada una de ellas debe valorar si “utiliza o prohíbe determinados usos”.
También los gobiernos de las universidades consideran que recae en el profesorado la tarea de comprobar si el alumno ha copiado. “Conocen a los estudiantes, los ven en clase, tutorías, seminarios…”, indica el comisionado rector para las TIC en la UAB, Jordi Hernández. En las universidades a distancia esto es más difícil, apunta.
Ante la irrupción de la IA, las universidades catalanas que son -algunas de ellas- las que han sido consultadas por este diario no van a reaccionar en caliente. “Es indiscutible que la IA va a cambiar la forma en que enseñamos y en que evaluamos a corto plazo”, asiente Manuel Jiménez, vicerrector de la UPF, un centro inmerso en un proceso de reflexión sobre el aprendizaje. «Pero esto requiere una reflexión profunda y no vamos a tomar una decisión precipitada».
«Está claro que la transmisión clásica de conocimiento del docente a los alumnos tal y como la conocemos ha caducado”, manifiesta Joan Gispets, vicerrector de la UPC, centro que ha encargado un estudio específico sobre este tema. La Politècnica de Catalunya también ha abierto un grupo de reflexión en el que durante 3 años se debatirá sobre el tipo de aprendizajes que deben recibir los estudiantes en el futuro. “Pero estoy seguro que la IA acabará siendo una aliada del profesor para enriquecer sus clases y del alumno para mejorar su aprendizaje” .

Ambiente de estudiantes universitarios en la biblioteca del edificio histórico de la UB en plaza Universidad
Ana Jiménez
“El cambio es profundo y no solo es conceptual, es también pedagógico, organizativo y económico”, indica Jiménez quien asegura que el bot se incorporará como una herramienta de apoyo muy valiosa y acabará naturalizándose en la clase, como los portátiles en las aulas. Se impulsará, considera, la personalización del aprendizaje, la creación original del conocimiento y se potenciará la presencialidad.
Hernández, comisionado para las TIC de la Autònoma de Barcelona cree -como experto, y no es una opinión que represente a la institución- que los movimientos defensivos de las universidades australianas o de las escuelas de Nueva York no van a servir de nada. «Los profesores del futuro tendrán que saber preguntar, preparar diseños pedagógicos diferentes, exigir a partir del nivel de ChatGPT… la solución no es poner puertas al campo sino adaptarse a la nueva realidad».
A su juicio estamos al inicio de «algo muy grande», una herramienta que ahora es imperfecta pero que, de momento, es gratis, de acceso al público generalista, que mejorará. Y profundizará en la información que maneja sobre derecho, medicina, ingeniería… «y vendrán otras herramientas».
La profesora del IES Vicens Vives, Ester Robles, está de acuerdo. «Llevo tres décadas en la enseñanza y en cada generación ha habido programas y aplicaciones que ha obligado al profesorado a replantearse las situaciones de aprendizaje y luchar contra el plagio», argumenta.
Alberto Romero, analista tecnológico y autor de The Algorithmic Bridge , una newsletter especializada en Inteligencia Artificial, apunta que la nueva realidad se sustenta en que “estos sistemas no copian de internet o de una base de datos. Modifican los textos por dentro y luego escriben de acuerdo con el contexto que se le ha puesto. Así que prácticamente nunca van a coincidir con textos ya existentes. La IA siempre está creando cosas nuevas”.
Los métodos utilizados hasta ahora para detectar si el trabajo de un alumno procedía de webs como El rincón del vago –que cotejaban los textos, como Turnitin– dejan de tener sentido.
Romero explica que estos métodos de prevención del plagio “no funcionan con la IA, porque están entrenados para distinguir a posteriori de la elaboración del texto”. “No es fiable –añade–. ChatGPT es suficientemente más avanzado, porque realmente sí que hay una parte de creatividad, aunque sea limitada”.
Lo que hace la IA de OpenAI , señala Romero, es crear textos “con solvencia, que no necesariamente van a estar bien, pero la estructura, te la van a dar en segundos y pueden hacer una redacción con una calidad universitaria”.
La propia OpenAI ha explicado que está ensayando un sistema que puede detectar si un texto procede de su IA. Se trata de una especie de “marca de agua” virtual que consiste en que la máquina, al crear un texto, utilice una serie de patrones en la distribución de palabras, que mantendrán una apariencia indistinguible para un humano, pero que revelarán su origen al ser analizada con una herramienta adecuada.
La compañía asegura que este sistema funciona bastante bien y afirma que “unos cientos de palabras parecen ser suficientes para obtener una señal razonable de que, sí, este texto procede de GPT”.
Alberto Romero advierte que, aunque este sistema funcione, puede ocurrir que otras IA de código abierto lleguen al mismo nivel de competencia que ChatGPT y la “marca de agua”, en este caso, no será extrapolable.
Existen ya algunas webs que prometen distinguir un texto humano de uno creado por una IA. En una prueba realizada con uno de los más famosos, indicó que un texto de ChatGPT era 99,99% humano. Parece que es demasiado pronto para estas soluciones.
Nerea Luis, responsable de IA en la empresa Sngular, observa que Microsoft ha hecho una fuerte inversión en OpenAI y tiene pactados unos derechos de explotación. “Por eso no hemos visto a ninguna compañía integrar este sistema”. “La hoja de ruta –indica– pasa por Microsoft Office. Es en ese momento cuando piensas que el debate de si se puede usar la IA en la educación está perdido”. “Ya podemos ir pensando en cómo adaptamos los tipos de ejercicios en la educación”, concluye.
Para Nerea Luis, recurrir a la máquina para hacer los trabajos académicos, “desde el punto de vista del estudiante, es feo”, aunque distingue aspectos positivos: “por otro lado están usando una de las herramientas que posiblemente tendrán que usar en el futuro. Esa interacción con el modelo de IA va a estar marcada en la vida de mucha gente”. “Es difícil prohibirla cuando ya tienes la tecnología”, apostilla.
La oleada que pueden provocar modelos de inteligencia artificial como el de OpenAI y otros que llegarán más adelante pueden transformar la educación para siempre. Ya está en la calle y en mano de una cantidad de alumnos que quizás todavía no es mayoritaria, pero también un día Google fue un buscador extraño que unos pocos empezaron a usar porque encontraba muy bien la información. Abróchense los cinturones.
Hemos entrevistado a GPT-3 sobre cómo será la vida en 2030 y esto es lo que nos ha dicho de Marte y el futuro (y dice que se llama Luis)
Representación de ChatGPT como un androide hecha por la IA Dall-E de OpenAI
REDACCIÓN
Alberto Romero, analista tecnológico y autor de The Algorithmic Bridge , una newsletter especializada en Inteligencia Artificial
Vicens Vives, Ester Robles, está de acuerdo. «Llevo tres décadas en la enseñanza y en cada generación ha habido programas y aplicaciones que ha obligado al profesorado a replantearse las situaciones de aprendizaje y luchar contra el plagio», argumenta.
Hernández, comisionado para las TIC de la Autònoma de Barcelona Los profesores del futuro tendrán que saber preguntar, preparar diseños pedagógicos diferentes, exigir a partir del nivel de ChatGPT… la solución no es poner puertas al campo sino adaptarse a la nueva realidad».




 Shutterstock
Shutterstock






 Anticuerpo monoclonal contra Coronavirus.
Anticuerpo monoclonal contra Coronavirus.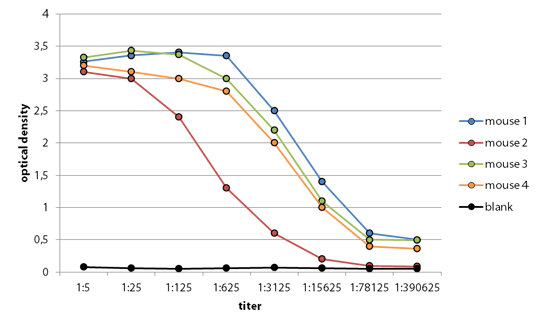 Desde que se aprobó rituximab, muchos otros se han incorporado a los tratamientos oncológicos (medicamentos como bevacizumab, trastuzumab y cetuximab, por citar algunos). Más recientemente, se han sumado a esta familia de ‘mAbs’ (del inglés monoclonal antibody), moléculas dirigidas a los puntos de control (los inhibidores de checkpoints pembrolizumab y nivolumab, sin ir más lejos) que frenan la respuesta del sistema inmunitario ante la célula tumoral, inaugurando una nueva era en la inmunoterapia contra el cáncer.
Desde que se aprobó rituximab, muchos otros se han incorporado a los tratamientos oncológicos (medicamentos como bevacizumab, trastuzumab y cetuximab, por citar algunos). Más recientemente, se han sumado a esta familia de ‘mAbs’ (del inglés monoclonal antibody), moléculas dirigidas a los puntos de control (los inhibidores de checkpoints pembrolizumab y nivolumab, sin ir más lejos) que frenan la respuesta del sistema inmunitario ante la célula tumoral, inaugurando una nueva era en la inmunoterapia contra el cáncer. 
 terapéutica que tiene un mecanismo de acción similar a la terapia CAR T en el sentido en que redirige el linfocito T contra la célula tumoral”, indica la hematóloga de la CUN. “La ventaja principal es que no requiere ningún proceso de producción, lo que hace que sea una terapia que está disponible inmediatamente y se pueda emplear en pacientes que no pueden esperar a la terapia CAR T. Esos resultados demuestran que el tratamiento es una nueva alternativa eficaz y segura en pacientes que han agotado los tratamientos disponibles. Hay varios estudios randomizados en marcha buscando confirmar estos resultados en otras poblaciones de pacientes con enfermedad menos avanzada”.
terapéutica que tiene un mecanismo de acción similar a la terapia CAR T en el sentido en que redirige el linfocito T contra la célula tumoral”, indica la hematóloga de la CUN. “La ventaja principal es que no requiere ningún proceso de producción, lo que hace que sea una terapia que está disponible inmediatamente y se pueda emplear en pacientes que no pueden esperar a la terapia CAR T. Esos resultados demuestran que el tratamiento es una nueva alternativa eficaz y segura en pacientes que han agotado los tratamientos disponibles. Hay varios estudios randomizados en marcha buscando confirmar estos resultados en otras poblaciones de pacientes con enfermedad menos avanzada”.

 Dios» con la letra de Einstein.
Dios» con la letra de Einstein. La primera hoja de la «Carta de Dios» de Albert Einstein.
La primera hoja de la «Carta de Dios» de Albert Einstein.