DAVID BAKER HA CREADO PROTEÍNAS
Y REVELADO LA ESTRUCTURA DE LAS EXISTENTES

LAS PROTEÍNAS Y SUS FUNCIONES.
Las proteínas transportan moléculas por todo el organismo, transmiten señales de una célula a otra y ayudan a defendernos de los agentes infecciosos. Existen miles de proteínas en nuestro cuerpo y cada proteína es única en su estructura y función. 
Nuestro ADN fabrica 20.000 tipos de proteínas diferentes, aunque la realidad es que seguramente haya muchas más.
La inmensa mayoría de nuestras proteínas son desconocidas
De las proteínas que conocemos , algunas tienen una función esencial .
Mencionaremos cinco de estas proteínas.
Tiene valores especiales la “p53”, llamada el guardian del genoma.
Es imprecindible en la lucha contra el cancer
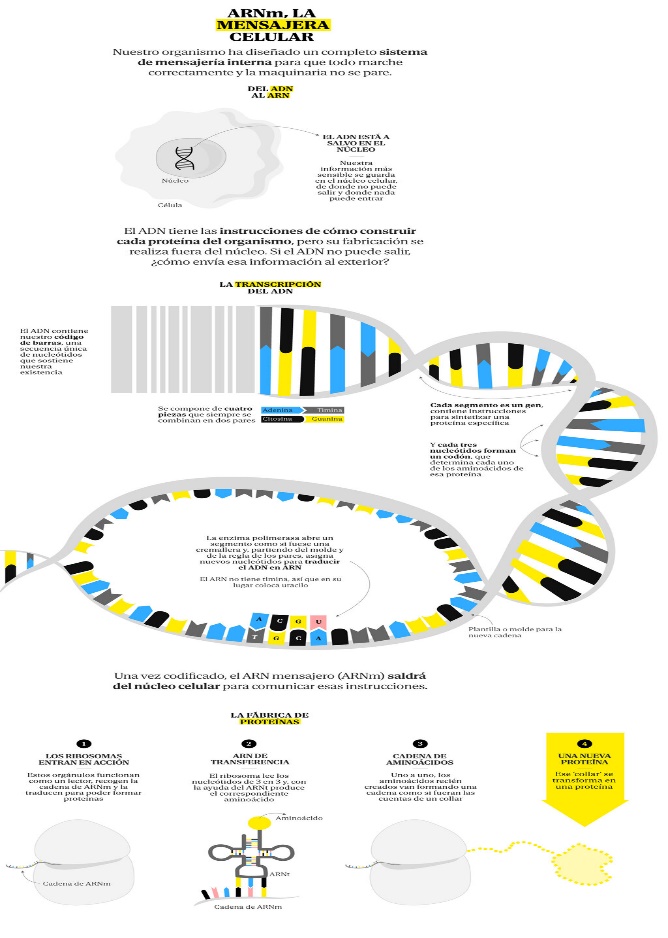
Para que una célula se divida y dé lugar a una nueva célula, necesita duplicar todo su contenido y luego partirse en dos. Por tanto, entre otras cosas, la célula necesitará hacer una copia de todo su ADN, que contiene las instrucciones para que la célula funcione, cada célula nueva que se forma necesita tener una copia completade esas instrucciones.
Este proceso de replicación del ADN, no siempre sale bien. Todos organismos sufren cambios accidentales en el ADN, ya sea debido al propio metabolismo de la célula o a causas ambientales incontrolables. Estos cambios del ADN son las mutaciones.
Si el ADN se altera, es muy posible que se produzcan consecuencias tanto para esa célula como para el individuo.
Las células, tienen una manera de evitar esto: la proteína p53.
La p53 es una proteína la “guardiana del genoma”.
Cuando una célula se está dividiendo y se produce una alteración accidental en su ADN, por ejemplo una mutación mientras se replicaba, la p53 frena la división, DETIENE TODA LA MAQUINARIA, dando tiempo a que se corrija el error. Si el error se corrige: se reinicia el ciclo; PERO si ese error no puede repararse, p53 induce la muerte de la célula para impedir que ese error se propague y tenga consecuencias más graves. Si la p53 no actuase ante las lesiones en el ADN, la célula iría transmitiéndo sus defectos y con el tiempo acumularían daños y mutaciones que podrían dar lugar a una célula cancerosa. 
La replicación del ADN no es la única manera en la que este puede dañarse: cosas como la exposición al humo del tabaco o la radiación ultravioleta del sol también pueden dañarlo. Y ahí p53 también actúa, intentando que se repare el daño, y, si el daño es tan grave que no puede repararse, condenando a la célula a morir por el bien del organismo. 
Las Kinesinas . Este tipo de proteínas y se encargan de transportar todo tipo de “paquetes” dentro de una célula, ya sean vesículas que contengan moléculas en su interior, cromosomas y hasta orgánulos enteros como las mitocondrias. Para ello, las kinesinas se desplazan sobre una especie de “carreteras” que atraviesan la célula, los llamados microtúbulos, que se distribuyen a lo largo de la célula tanto para darle forma y estructura como para el transporte en su interior.
La transmisión de partículas o de señales en general es común en todas las células.
Una neurona libera un tipo de neurotransmisor (por ejemplo: dopamina, adrenalina o serotonina); este viaja a través del espacio entre una neurona y otra, llamado sinapsis, y es captado por los receptores de la siguiente neurona, transmitiendo así la señal de una neurona a la siguiente
Los neurotransmisores se sintetizan en el cuerpo de la neurona, pero para poder transmitir el impulso nervioso necesitan ser transportados hasta la otra parte de la neurona. Y aquí es donde entran en juego las kinesinas, utilizando los microtúbulos que recorren el interior de las neuronas, transportan vesículas cargadas de neurotransmisores. De esta forma, los neurotransmisores llegan a su destino, el extremo de la neurona, listos para que, cuando llegue el impulso eléctrico, puedan liberarse y transmitir la señal hacia la siguiente neurona.
“El Colágeno”
Las proteínas como moléculas van de un lugar para el otro, y transportan cosas, y digieren alimentos y transforman moléculas aquí y allá, pero algunas proteínas son un tanto distintas. Algunas proteínas sencillamente conforman nuestra estructura, dando lugar a nuestra piel, nuestros huesos, cartílago y ligamentos, o incluso el recubrimiento de nuestros órganos. Una de las proteínas estructurales más importantes que conocemos: el Colágeno es una proteína dura y fibrosa que constituye básicamente un tercio de las proteína del cuerpo humano. Las moléculas de colágeno suelen agruparse para formar fibrillas largas y delgadas que actúan como estructuras de soporte y anclan las células entre sí, proporcionando, entre otras cosas, resistencia a los tendones y ligamentos o elasticidad a la piel.
Existen muchos tipos de colágeno que forman las diferentes estructuras de nuestro cuerpo. Los más abundantes son estos:
Colágeno I: se encuentra en huesos, tendones y órganos.
Colágeno II: se encuentra principalmente en el cartílago.
Colágeno III: forma una malla de soporte que recubre y sostiene algunos de nuestros órganos y tejidos, como el hígado, las paredes de los vasos sanguíneos o la dermis de la piel.
Colágeno IV: forma parte de la red de soporte de algunas de nuestras células, manteniéndolas en su sitio.
Colágeno V- que se encuentra en el cabello y las uñas, entre otras cosas.
Existen muchos tipos de trastornos asociados con el colágeno, como por ejemplo el síndrome de Ehlers-Danlos, en el que el colágeno no se forma como es debido y los afectados presentan afecciones como una hiper elasticidad de la piel, hipermovilidad de las articulaciones, mala cicatrización y fragilidad de sus vasos sanguíneos, además de muchísimas otros problemas. LAS PROTEÍNAS Y SUS FUNCIONES.
Las proteínas transportan moléculas por todo el organismo, transmiten señales de una célula a otra y ayudan a defendernos de los agentes infecciosos. Existen miles de proteínas en nuestro cuerpo y cada proteína es única en su estructura y función. 
Nuestro ADN fabrica 20.000 tipos de proteínas diferentes, aunque la realidad es que seguramente haya muchas más.
La inmensa mayoría de nuestras proteínas son desconocidas
De las proteínas que conocemos , algunas tienen una función esencial .
Dada su abundancia, dedicare solo a cinco de estas proteínas.
Antes que nada, este tema fue elegido por los Patreons

Patreon es una plataforma de suscripción que facilita que los creadores obtengan ingresos.
La primera proteina con valores extraordinarios ess la “p53”, llamada el guardan del genoma. Es imprecindible en la lucha contra el .
cáncer.
Nuestras células se dividen constantemente, para regenerar los tejidos en unas condiciones de equilibrio entre células que mueren y nuevas célulasque se dividen. Que una célula se divida para dar lugar a una nueva célula es un proceso muy complejo.
Para que una célula se divida y dé lugar a una nueva célula, necesita duplicar todo su contenido y luego partirse en dos. Por tanto, entre otras cosas, la célula necesitará hacer una copia de todo su ADN. El ADN es esa molécula que contiene las instrucciones para que la célula funcione, cada célula nueva que se forma necesita tener una copia completade esas instrucciones.
El problema es que este proceso de replicación del ADN, no siempre sale bien.
Todos organismos sufre cambios accidentales en el ADN,
ya sea debido al propio metabolismo de la célula o a causas ambientales que no podemos controlar. Los cambios en nuestro ADN son las mutaciones.
Si el ADN se alter, es muy posible que se produzcan consecuencias tanto para esa célula como para el individuo.
Las células, tienen una manera de evitar esto: la proteína p53.
La p53 es una proteína la “guardiana del genoma”.
Cuando una célula se está dividiendo y se produce una alteración accidental en su ADN, por ejemplo una mutación mientras se replicaba, la p53 frena la división, DETIENE TODA LA MAQUINARIA, dando tiempo a que se corrija el error. Si el error se corrige: se reinicia el ciclo; PERO si ese error no puede repararse, p53 induce la muerte de la célula para impedir que ese error se propague y tenga consecuencias más graves. Si la p53 no actuase ante las lesiones en elADN, la célula iría transmitiéndolas sus defectos y con el tiempo acumularían daños y mutaciones que podrían dar lugar a una célula cancerosa. 
De hecho, la replicación del ADN no es la única manera en la que este puede dañarse: cosas como la exposición al humo del tabaco o la radiación ultravioleta del sol también pueden dañarlo. Y ahí p53 también actúa, intentando que se repar el daño, y, si el daño es tan grave que no puede repararse, condenando a la célula a morir por el bien del organismo. 
La Kinesina .Este tipo de proteínas son lo que llamamos kinesinas, y se encargan de transportar todo tipo de “paquetes” dentro de una célula, ya sean vesículas que contengan moléculas en su interior, cromosomas y hasta orgánulos enteros como las mitocondrias. Para ello, las kinesinas se desplazan sobre una especie de “carreteras” que atraviesan la célula, los llamados microtúbulos, que son algo así como una red de autopistas que se distribuyen a lo largo de la célula tanto para darle forma y estructura como para el transporte en su interior.
La transmisión de partículas o de señales en general es común en todas las células.
Las neuronas son las células por excelencia de nuestro sistema nervioso, y como tal, son las encargadas de transmitir el impulso nervioso. Y transmiten impulso nervioso de unas a otras a través de unas moléculas llamadas neurotransmisores.
Una neurona libera un tipo de neurotransmisor (por ejemplo: dopamina, adrenalina o serotonina); este viaja a través del espacio entre una neurona y otra, llamado sinapsis, y es captado por los receptores de la siguiente neurona, transmitiendo así la señal de una neurona a la siguiente
Los neurotransmisores se sintetizan en el cuerpo de la neurona, aquí, pero para poder transmitir el impulso nervioso, necesitan ser transportados hasta la otra punta de la neurona. Y aquí es donde entran en juego las kinesinas, utilizando los microtúbulos que recorren el interior de las neuronas, transportan vesículas cargadas de neurotransmisores. De esta forma, los neurotransmisores llegan a su destino, el extremo de la neurona, listos para que, cuando llegue el impulso eléctrico, puedan liberarse y transmitir la señal hacia la siguiente neurona.
La siguiente proteína es una de las más populares tanto por su implicación en la industria cosmética como por la enorme cantidad de productos que se venden relacionados con ella: “El Colágeno”
Las proteínas como moléculas van de un lugar para el otro, y transportan cosas, y digieren alimentos y transforman moléculas aquí y allá, pero algunas proteínas son un tanto distintas. Algunas proteínas sencillamente conforman nuestra estructura, dando lugar a nuestra piel, nuestros huesos, cartílago y ligamentos, o incluso el recubrimiento de nuestros órganos.
Nuestro cuerpo, por muy activo que sea, no tendría sentido sin una estructura que le diese forma y consistencia.
Una de las proteínas estructurales más importantes que conocemos: el Colágeno es una proteína dura y fibrosa que constituye básicamente un tercio de las proteína del cuerpo humano. Las moléculas de colágeno suelen agruparse para formar fibrillas largas y delgadas que actúan como estructuras de soporte y anclan las células entre sí, proporcionando, entre otras cosas, resistencia a los tendones y ligamentos o elasticidad a la piel.
Existen muchos tipos de colágeno que forman las diferentes estructuras de nuestro cuerpo. Los más abundantes son estos:
Colágeno I: se encuentra en huesos, tendones y órganos.
Colágeno II: se encuentra principalmente en el cartílago.
Colágeno III: forma una malla de soporte que recubre y sostiene algunos de nuestros órganos y tejidos, como el hígado, las paredes de los vasos sanguíneos o la dermis de la piel.
Colágeno IV: forma parte de la red de soporte de algunas de nuestras células, manteniéndolas en su sitio.
Colágeno V- que se encuentra en el cabello y las uñas, entre otras cosas.
Una manera de entender la importancia que tiene una proteína en nuestro cuerpo, es observar qué ocurre en aquellas enfermedades en las que esa proteína no funciona correctamente.
Existen muchos tipos de trastornos asociados con el colágeno, como por ejemplo el síndrome de Ehlers-Danlos, en el que el colágeno no se forma como es debido y los afectados presentan afecciones como una hiper elasticidad de la piel, hipermovilidad de las articulaciones, mala cicatrización y fragilidad de sus vasos sanguíneos, además de muchísimas otros problemas.
El colágeno es una proteína fundamental para dar estructura a nuestro cuerpo .
Serotonina. conocida como “la hormona de la felicidad”.
La serotonina calibra nuestras necesidades básicas: es decir, cuando nuestros niveles de comida, agua y sueño son suficientes, tendríamos niveles más altos de serotonina, lo que se correlacionaría con una mayor percepción de felicidad; sin embargo, cuando faltan estas necesidades fisiológicas, se produce un efecto adverso. La serotonina es una hormona compleja y sus funciones en nuestro sistema nervioso central son muy amplias:
Ayudan a regular la temperatura, el apetito, memoria, aprendizaje, los ciclos del sueño, además del comportamiento y estado de ánimo. Tanto es así que los niveles bajos de esta hormona a menudo se asocian con trastornos como la depresión, la ansiedad o el trastorno obsesivo-compulsivo. De hecho, algunos de los fármacos que suelen utilizarse para tratar estos trastornos incrementan la actividad de la serotonina en el cerebro. Aunque sigue habiendo discusión respecto a si la serotonina debería ser la única “diana” en este tipo de tratamientos, ya que son trastornos realmente complejos y de los que todavía nos queda muchísimo por conocer.
A pesar de que la serotonina se asocie popularmente más al cerebro, lo cierto es que se estima que el 90% de la serotonina en el cuerpo humano no está en el cerebro, sino que se almacena en unas células especializadas de nuestros intestinos. Cuando estas células del intestino detectan que hay comida en el estómago, liberan serotonina como respuesta, que estimula los movimientos intestinales para ayudar con la digestión y reduce el apetito. La serotonina juega un papel en la coagulación de la sangre y contribuye a que las plaquetas de nuestra sangre se agreguen formando un coágulo y a que nuestros vasos se estrechen, lo cual es importante a la hora de curar las heridas que podamos hacernos.
Los anticuerpos son unas de las proteínas más populares en la actualidad: “vacunas contra el COVID, tests de anticuerpos, inmunización!
Los anticuerpos, también conocidos como inmunoglobulinas, son unas proteínas con forma de tirachinas que secretan nuestras células inmunitarias. El objetivo es que estos anticuerpos se unan a los agentes infecciosos y los “marquen” para que otras células inmunitarias los localicen y eliminen más fácilmente. Los anticuerpos se liberan cuando tenemos una infección para que esta se elimine lo antes posible y no cause grandes daños. Pero no solo sirven para actuar en el momento de la infección, sino que esconden una de las claves que hace que nuestro sistema de defensa sea tan increíble:
La memoria inmunitaria. Nuestro sistema inmunitario no solo es capaz de detener una infección en el momento, sino también de “guardar información” sobre el microorganismo que la causó, por si acaso nos lo topamos en un futuro. Esto se consigue gracias a las llamadas células de memoria, que dan vueltas por la sangre y que, si se encontrasen de nuevo con el microorganismo, secretarían rápidamente anticuerpos específicos contra él, dando lugar a una respuesta inmunitaria mucho más rápida y masiva que la primera. Esta es la clave de cómo funcionan las vacunas: algunas vacunas, por ejemplo, están formadas por microorganismos atenuados o inactivados, o directamente por partes de los mismos. Esto hace que las vacunas actúen como una especie de “simulacro de infección”: ponen en contacto a nuestro sistema inmunitario con el microorganismo en cuestión para que genere células de memoria y por tanto quede “entrenado” para una posible infección en un futuro.
El colágeno es una proteína fundamental para dar estructura a nuestro cuerpo .
La siguiente proteína es una hormona, popularmente conocida como la hormona de la felicidad.
Serotonina.
La serotonina es conocida como “la hormona de la felicidad”. podríamos decir, que la serotonina nos ayudaría a calibrar lo satisfechas que están nuestras necesidades básicas: es decir, cuando nuestros niveles de comida, agua y sueño son suficientes, tendríamos niveles más altos de serotonina, lo que se correlacionaría con una mayor percepción de felicidad; sin embargo, cuando faltan estas necesidades fisiológicas, se produce un efecto adverso. Pero la serotonina es una hormona más compleja que eso, ya que sus funciones en nuestro sistema nervioso central son muy amplias: entre otras cosas, ayudan a regular la temperatura, el apetito, memoria, aprendizaje, los ciclos del sueño, además del comportamiento y estado de ánimo. Tanto es así que los niveles bajos de esta hormona a menudo se asocian con trastornos como la depresión, la ansiedad o el trastorno obsesivo-compulsivo. De hecho, algunos de los fármacos que suelen utilizarse para tratar estos trastornos incrementan la actividad de la serotonina en el cerebro.. 
El 90% de la serotonina en el cuerpo humano no está en el cerebro, sino que se almacena en unas células especializadas de nuestros intestinos y se libera como respuesta, que estimula los movimientos intestinales para ayudar con la digestión y reduce el apetito. Pero no solo eso, sino que la serotonina juega un papel en la coagulación de la sangre. La serotonina contribuye a que las plaquetas de nuestra sangre se agreguen formando un coágulo y a que nuestros vasos se estrechen, lo cual es importante a la hora de curar las heridas que podamos hacernos.
Los anticuerpos son unas de las proteínas más populares en la actualidad: “vacunas contra el COVID, tests de anticuerpos, que si inmunización! Los anticuerpos, también conocidos como inmunoglobulinas, estos anticuerpos se unen a los agentes infecciosos y los “marcan” para que otras células inmunitarias los localicen y eliminen.
La memoria inmunitaria. Nuestro sistema inmunitario no solo es capaz de detener una infección en el momento, sino también de “guardar información” sobre el microorganismo que la causó, por si acaso nos lo topamos en un futuro. Esto se consigue gracias a las llamadas células de memoria, que dan vueltas por la sangre y que, si se encontrasen de nuevo con el microorganismo, secretarían rápidamente anticuerpos específicos contra él, dando lugar a una respuesta inmunitaria mucho más rápida y masiva que la primera. Esta es la clave de cómo funcionan las vacunas: algunas vacunas, por ejemplo, están formadas por microorganismos atenuados o inactivados, o directamente por partes de los mismos. Esto hace que las vacunas actúen como una especie de “simulacro de infección”: ponen en contacto a nuestro sistema inmunitario con el microorganismo en cuestión para que genere células de memoria y por tanto quede “entrenado” para una posible infección en un futuro.
DAVID BAKER HA CREADO PROTEÍNAS Y REVELADO LA ESTRUCTURA DE LAS EXISTENTES
Es candidato al Nobel de Medicina?
Ha diseñado proteínas y van a revolucionar la medicina, y la biología.
Hemos desarrollado herramientas para conocer con precisión sin precedentes la estructura tridimensional de las proteínas de animales, plantas y bacterias; y en solo unos días, cuando antes se requerían años para tan solo aproximarse a ese conocimiento.
Las proteínas son las partículas de la vida: ¿poder crearlas es crear vida?
Es una buena analogía, porque las proteínas están implicadas en todo: de capturar la energía del sol a la neurodegeneración causada por el Alzheimer o en el diseño de vacunas y las estructuras de los virus. Las proteínas lo hacen todo y poder crearlas es clave.
¿Y conocer su estructura e incluso diseñarlas nuevo ya tiene aplicaciones médicas?
Ya se está inyectando una vacuna coreana contra la COVID, diseñada con nuestra nueva herramienta de inteligencia artificial, el
RoseTTAFold, y un spray nasal que también vacuna contra otros virus respiratorios, como la gripe y otras terapias para las células cancerígenas o el parásito de la malaria; pero son incluso más importantes las puertas que abre ese nuevo conocimiento…
¿Cuál es la más relevante?
La mayoría de los medicamentos actuales se fabrican introduciendo modificaciones en las proteínas que ya existen.
¿Ahora diseñarán proteínas para curar?
Ahora diseñaremos proteínas nuevas para medicamentos sofisticados y mejorados, más precisos y robustos que traten el cáncer sin efectos secundarios o para fabricar vacunas rápidas contra nuevas pandemias.
¿En qué trabajan exactamente ahora?
Me fascina la posibilidad de diseñar proteínas que bloquearan la acumulación de amiloides que causan el Alzheimer. Y más allá de la biomedicina: ¿por qué no crear proteínas que rompan los plásticos en el océano?
¿Por qué hemos tardado tanto en poder fabricar proteínas?
Había dos problemas difíciles de resolver: las proteínas son responsables de múltiples funciones en nuestros organismos y en todos los vivientes –y hay billones en la naturaleza– y lo que hacen depende de sus secuencias de aminoácidos…
Los genes que forman nuestro ADN están compuestos de proteínas en secuencias de A (adenina), T (timina), G (guanina) y C (cytosina) y ese era el primer problema: averiguar su estructura en cada gen; el segundo era diseñarlas. Y hemos resuelto los dos.
Además ha fundado varias startups biomédicas para fabricar medicamentos.
Y hemos logrado llegar hasta los ensayos clínicos en humanos contra las enfermedades inflamatorias, como las intolerancias celíacas; pero, sobre todo, me interesan las relacionadas con las enfermedades del envejecimiento, que cada día son más y más extendidas. Y crear proteínas cambia el juego.
¿Por qué le interesan las degenerativas?
Porque los medicamentos que estamos diseñando a partir de las proteínas que creamos son más eficientes y baratos que los de anticuerpos, que son los más usados hoy. Y además, podemos diseñar nuevas proteínas ad hoc para otros usos revolucionarios.
Planeamos nuevos sistemas de sintetizar la energía solar creando proteínas al efecto; y hay otros materiales biológicos muy interesantes con los que podríamos trabajar ahora, como el hueso, las conchas de moluscos o los dientes, que interactúan en la naturaleza con químicas inorgánicas como el calcio y los carbonatos. Abren campos fascinantes.
¿Y el envejecimiento no es una cuestión también de proteínas e inflamación?
Lo que hacemos ahora en el laboratorio es diseñar genes sintéticos con las proteínas que hemos creado y luego comprobamos si actúan como habíamos previsto al diseñarlas para frenar un proceso degenerativo.
¿Y funciona?
Lo que ya es seguro es que ese proceso se ha acortado sustancialmente y eso nos da una enorme ventaja para el diseño de vacunas contra nuevas pandemias. Hoy podríamos diseñar la vacuna COVID en solo dos semanas.
¿Por qué ya somos tan rápidos?
Porque aplicamos la inteligencia artificial del RoseTTAFold. Pero el envejecimiento es un proceso complejo sobre el que apenas tenemos experiencia estadística: nunca tantos habían vivido tanto. Ahora necesitamos más hipótesis centrales de trabajo.
Y nosotros, a ustedes.
Tengo un gran equipo en el que destacan algunos científicos españoles brillantes. Por cierto: ¿la dieta mediterránea justifica la longevidad española por sí sola?
Tenemos un buen sistema de salud.
¡Ah! Eso es importante también.
Bibliografia
Instragram,Twitch o TikTok.













 Cifras de supervivencia de cáncer en 2023 y los factores de riesgo asociados. Infografía: SEOM.
Cifras de supervivencia de cáncer en 2023 y los factores de riesgo asociados. Infografía: SEOM.




 mas de la esquizofrenia y de trastorno mayor de ánimo tal como la depresión.
mas de la esquizofrenia y de trastorno mayor de ánimo tal como la depresión.