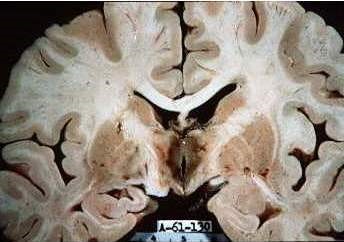
Los Egipcios consideraron al Cerebro como un órgano accesorio, y nunca lo estudiaron porque lo extraían por la nariz durante las practicas de embalsamarlo, Aristoteles ubicó los procesos mentales en el corazón, hubo que esperar mas de una centuria para que Hipócrates de Cos, Paracelso, pensaran en el cerebro como órgano del pensamiento y Juan de Huarte de San Juan que vivió entre 1529 y 1588, tuvo ya una idea clara de la función mental del cerebro, Descarte ya en el siglo XVII creía que la mente, la llamada alma, era ajena al cuerpo aunque se relacionaba a través de la glándula pineal con el cerebro, aunque su experiencia se relegaba solo a haber visto los dibujos de Vesalio.
Sorprende esta torpeza en los tiempos pasados donde los niveles de creación eran importantes, por lo menos en arte, aunque por supuesto no en biología. Cualquier hombre, de nuestro tiempo, que piense, ¿conque estoy pensando?, sabe de inmediato que lo hace con algo alojado en el cráneo. Esta pregunta la hago cientos de veces a personas de mi entorno y no falla ni una sola. No me resigno a admitir lo contrario. Me cuesta esfuerzo aceptar esta torpeza, y me explico parte de la estupidez humana.
Este órgano tiene la capacidad de convertir algo tan físico como la materia organica en pensamientos,como dice Morgado, solo el cerebro es capaz, aunque con mucho esfuerzo y tiempo, de resolver estas dudas.
Sorprende la multitud de células. El humano posee 10 elevado a11 neuronas con sus multiples ramificaciones y conexiones.
Muchos problemas mentales son automáticos e inconscientes sin embargo la mayoría de ellos como las percepciones, las emociones o la memoria también son conscientes aunque buena parte de sus componentes sea siempre inconsciente.
Se define la conciencia como un estado de la mente en la que nos damos cuenta de cosas que pasan en ella aunque no de todo lo que pasa. Y la perdemos, cuando, dormimos, nos anestesian o sufrios un traumatismo.
Nos permite la conciencia recordar o sentir miedo pero lo que no percibimos puede ser no bien recordado
La conciencia es un estado crítico de la mente y consiste en un pensamiento sobre otro pensamiento es decir su naturaleza es reflexiva
El mundo que nos rodea, lo percibimos y concienciamos, a través del cerebro, que es el órgano encargado de disminuir la incertidumbre del mundo que nos rodea
Conocemos el mundo por el cerebro, y la mente es su función principal.
El cerebro filtra la información que recibe, la procesa y la conciencia a su manera.
Por tanto, la realidad del mundo que percibimos esta personalizada. Solo lo que existe en nuestra mente nos es conocido, lo demás no existe.
Cuando el cerebro se altera, también lo hace la mente.
Los órganos de los sentidos, son los informadores del mundo que nos rodea, y tras su análisis es concienciado. Pero en los poderes de la mente, esta imaginar e imagina lo que no percibe, y es capaz de convertirlo en su realidad
La conciencia, se encarga de reconocer mi yo y mi entorno.
La gran pregunta de todos los tiempos y del actual es: Como la materia de nuestra biología se convierte en imaginación.
La mente identifica al hombre y a su vez nos identificamos con ella.
¿La mente nos ha sido añadida al cuerpo, pero es diferente de el ¿.
Siempre el binomio, cuerpo y alma, o simplemente un todo que funciona . Lo que si esta claro es que conocernos, nuestro cuerpo a través de algo que es invisibe a los sentidos y es nuestra mente.
Lo que percibimos esta modificado y es absolutamente irreal, nadie tiene una flor en el cerebro, pero la disfrutamos con realidad, pero es de verdad esta realidad’¿. Y además la mayor parte de los procesos mentales, son inconscientes y los procesos conscientes tienen una base inconsciente de cantidad de procesos, sino de todos.
De forma que esto es muy difícil de entender. Incluso cuando se afirma que el cerebro funciona de tal forma que acopla lo físico y mental. Esto lo sabe todo el mundo, pero como lo hace, no lo sabe nadie y nos entusiasma su estudio, eso sí lo intentamos y con repetición.
Cuándo el cerebro de los animales después de la fase reptiliana, desarrollaron el lóbulo limbico , su intervención complicó el resultado, la compresión de lo nuevo y los ambientes sociales, llevan a usar las emociones, como fijadoras de sentimientos que modifican lo percibido, la memoria, los sueños y multiples procesos mas, percibidos y subliminales.
Sobre la organicidad del tejido nervioso, del cerebro, la mente extrae del medio información, la modifica y la incorpora y complica el resultado de forma inimaginable y además confiere a este cerebro de forma progresiva, más y más capacidad. El medio, influencia, más que los cromosomas.
Solo existe en el cerebro, para poder medir, una energía electroquímica y mediante la actividad de las células nerviosas y con sus conexiones podemos percibir lo interno y externo a nuestra biología, que no tiene porque coincidir con la realidad, simplemente lo imaginamos
La idea que la mente esta en el cerebro y fuera de él, es apetitosa, y tranquiza a muchos. Pero la mente con nuestro medir, no es entendible, como materializa nuestras ideas. Quizas porque estamos hechos de la misma materia que nuestros sueños. Una forma romántica de ayudar a tanto desatino.
Despues de Descartes optamos por la teoría unicista, la mente es el producto de nuestro cerebro. No esta en región alguna y por tanto hasta ahora no se localiza , aunque algunas regiones cerebrales al lesionarse, modifican su función. La mente es una función y estas no se miden o aun no tenemos medios.
Dice Morgado, que las piernas sirven para caminar, pero la voluntad de hacerlo no esta en ellas.
Que hacemos los humanos para tener dentro de la cabeza el universo. El cerebro inventa la luz o los colores, que no existen fuera, como tampoco está fuera el dolor. Lo que percibimos en el cerebro, es una realidad virtual producto de la interpretación de las ondas electromagnéticas que nos llegan por los órganos de los sentidos
Podríamos definir el cerebro como una máquina predictiva encaminada a disminuir la incertidumbre del mundo que nos rodea
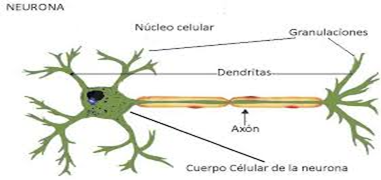
El tejido nervioso consta de células nerviosas o neuronas y de células de soporte o glía. La célula nerviosa, o neurona, es propiamente la unidad elemental básica del sistema nervioso. El encéfalo humano contiene 100.000 millones de neuronas junto con una variedad de células gliales que ayudan a sostener y mantener la integridad física y fisiológica de las neuronas. La neurona consta de cuerpo celular o pericarion, partiendo del cuerpo celular neuronal se observan múltiples ramificaciones o dendritas, que son prolongaciones cortas y cónicas que reciben los impulsos nerviosos aferentes. Cada neurona da lugar a un único axón, que transporta los impulsos desde el cuerpo celular y a través de las sinapsis, a otras partes del cerebro o médula espinal, a bordo de las dendritas.
El cerebro funciona con electricidad. Los impulsos eléctricos son transportados desde el axón terminal a dendritas de otras neuronas en zonas diana apropiadas del encéfalo
El cerebro dispone sus células en su parte externa en lo que denominamos corteza (córtex cerebral) y en acúmulos más profundos que constituyen los ganglios basales
Es en la corteza cerebral donde se sitúan las funciones más finas sensitivas, motoras, y psicológicas. Según su arquitectura, es decir del número de capas en que se disponen las neuronas, en la corteza cerebral, se pueden diferenciar, claramente dos zonas: por un lado el paleo y arquicórtex, formada por tres capas de neuronas y por otro, el neocórtex.
La organización de las conexiones del encéfalo permite que múltiples impulsos excitatorios e inhibitorios sean integrados en una única experiencia mental.
Zonas específicas de la corteza cerebral reciben las aferencias de partes concretas del organismo, mientras que el córtex calcarino está retinotópicamente organizado, y el córtex auditivo está tonotópicamente organizado. Cada región sensitiva primaria tiene conexiones con áreas de asociación de modalidad específica, donde tiene lugar la convergencia e integración de diferentes atributos de la experiencia sensorial. Los axones de diferentes áreas sensitivas de asociación de modalidad específica, empiezan a converger en lo que se denominan áreas de asociación multimodal. En estas áreas se ha demostrado que existen neuronas, que por ejemplo, se activan en respuesta al estímulo visual, encontrándose entremezcladas con neuronas que responden a estímulos auditivos, y con neuronas que responden a estímulos sensitivos múltiples. Es decir forman una red neuronal, donde probablemente, todo se conecta con todo.
Una de las características del homo sapiens es el peso de su cerebro, el espesor de la corteza cerebral, la amplitud del lóbulo frontal y el diámetro biparietal. El cerebro humano pesa 1300 gramos, mientras que el del chimpancé pesa sólo 350. No siempre el peso del cerebro es expresivo de la capacidad cerebral ya que la ballena los elefantes y otros animales tienen un mayor peso del cerebro así como un mayor volumen de este. La relación del volumen del cerebro comparado con el peso del animal es la proporción que marca la potencia intelectual de un mamífero.
Antonio Damasio experto en neurofisiología dice que la vida psíquica es el esfuerzo permanente entre dos cerebros un, cerebro emocional inconsciente preocupado sobre todo por sobrevivir y ante todo conectado al cuerpo. Otros cerebro cognitivo, consciente, racional y volcado en el mundo externo. Es todo cerebro son independientes entre cada uno de ello contribuye de forma diferente a nuestra experiencia de vida y a nuestro comportamiento.
El doctor Francisco Mora de una manera dramática dice “” como un montón de neuronas enmarañadas unas con otras pueden dar lugar a un a un individuo que piensa y siente, que llora y ríe y con ello levanta su mirada hacia el infinito universo y se pregunta por su existencia y su sentido”.
El estudio del cerebro sus funciones y su vida psíquica, están solo suavemente conocidas. Hace falta un gran esfuerzo y tiempo para entenderlas enteras.
El conocimiento parcial de las maquinas que funcionan con espíritu, no esta terminado y por supuesto el error esta asegurado casi siempre que vaticinamos. Solo lo que está en mi mente existe, pero para mi , no para los demas .
LAS CÉLULAS MADRE NEURALES DEL BULBO OLFATORIO GENERAN NEURONAS EN LA ETAPA ADULTA
La neurogénesis en el hombre es un problema muy discutido
Hasta ahora solo determinadas zonas del diencéfalo relacionadas con la memoria, parecía claro que generan nuevas neuronas. La zona subgranular del giro dentado del hipocampo y la zona subventricular
Neurona (verde) generada a partir de células madre neurales del bulbo olfatorio, con botones sinápticos formados por la proteína sinapsina (en rojo), lo que indica contactos sinápticos (C. Vicario et al.).
Un estudio coordinado por el Consejo Superior de Investigaciones Científicas (CSIC) ha identificado un nicho de células madre neurales (NSC, por sus siglas en inglés) en la zona central del bulbo olfatorio del cerebro. Estas células generan neuronas en un proceso llamado neurogénesis adulta. El hallazgo, que se publica en la revista Stem Cells, aumenta el conocimiento de la plasticidad estructural de esta región del cerebro.
La neurogénesis adulta, proceso mediante el que se generan nuevas neuronas en la edad adulta a partir de células madre neurales, no ocurre en todo el encéfalo. Hasta ahora se apuntaba solo a dos regiones: la zona subgranular del giro dentado del hipocampo y la zona subventricular.
“Aunque estudios previos ya sugerían la existencia de células madre neurales en el bulbo olfatorio, su localización, identidad y capacidad de generar neuronas in vivo ha sido poco explorada”, apunta Carlos Vicario, investigador del CSIC en el Instituto Cajal (IC-CSIC) y director del trabajo. “Nuestros resultados muestran, por primera vez, que se generan nuevas neuronas y que éstas se incorporan activamente a los circuitos sinápticos del bulbo olfatorio”.
Helena Mira, científica del CSIC en el Instituto de Biomedicina de Valencia (IBV-CSIC) y coautora del artículo, señala que “el bulbo olfatorio de los mamíferos es una estructura muy plástica que juega un papel fundamental en la transmisión de información olfativa. Esta plasticidad se basa, en gran medida, en la neurogénesis que tiene lugar en la zona subventricular y, como mostramos en nuestro estudio, también podría depender de las células madre neurales locales del bulbo olfatorio”.
Los investigadores han podido marcar las células madre neurales inyectando partículas retrovirales portadoras de la proteína verde fluorescente (GFP, por sus siglas en inglés) en el bulbo olfatorio. “Gracias a la proteína GFP, y a través de la microscopía confocal y técnicas de análisis de imagen, no solo hemos podido ver la formación de neuronas a partir de dichas células, sino que hemos seguido y analizado la transición desde la célula madre neural a la neurona inmadura, y de esta a la neurona madura. La colaboración y experiencia en microscopía electrónica de Carlos Crespo, de la Universidad de Valencia, ha sido fundamental para demostrar que las nuevas neuronas establecen sinapsis”, comenta Vicario.
“Dado que no se ha detectado neurogénesis en el bulbo olfatorio humano en la totalidad de estudios que se han realizado hasta la fecha, no se puede descartar que este proceso tenga lugar en unas circunstancias determinadas y pueda estar condicionado bien por la estimulación olfativa o la respuesta a un daño producido por una lesión o una infección, como puede ser la provocada por el coronavirus SARS-CoV-2, que afecta al olfato”, concluye el investigador.
Este estudio parece claro. La neurogénesis además de aparecer en circuitos de la memoria, se producen también en el bulbo olfatorio.
Los autores, con precaución admiten la posibilidad que determinadas condiciones especiales den lugar a esta neurogénesis, de nuevo el coronavirus puede hacer cosas distintas a las conocidas
No obstante es positivo que se puedan encontrar nuevos lugares de neurogénesis, y aunque sean producto de las circunstancias, como el Coronavirus, abren la posibilidad de que de alguna manera se puede inducir neurogénesis.
Como siempre hacen falta mas trabajos que repitan el hallazgo. Y esperara que desaparezca el coronavirus y ver si se repite el hallazgo
Bibliografía
Stem Cells, Consejo Superior de Investigaciones Científicas (CSIC)
Se parecen en algo estas dos maquinas, una biológica y otra mecánica?.
Intento buscar similitudes, que las tienen sin duda, pero solo hasta llegar a las funciones superiores del cerebro, donde se germina lo espiritual. Esto hasta ahora no se encuentra, pero si otras muchas cualidades
¿Cómo es y cómo funciona nuestro cerebro?
El cerebro está, implicado en todas las funciones que nos mantienen vivos.
Sirve para pensar, razonar Y comunicarnos, soportan nuestra homeostasis y nos permite también para cosas tan básicas como respirar o parpadear, así como para soñar, y emocionarnos.
A través de los sentidos, el cerebro recibe un flujo enorme de información del mundo que nos rodea, la procesa y hace que cobre significado, organiza y controla el movimiento.
Un cerebro adulto pesa entre 1.300 y 1.400 gramos. Contiene unos 100.000 millones de neuronas y una cantidad mucho mayor de sinapsis, que permiten la conexión entre neuronas.
El cerebro se encarga de mantener tantos las funciones mas elemantales como mantener un equilibrio orgánico y químico, como para soñar y adivinar el futuro..
El encéfalo consta de tres grandes áreas: el PROPIO CEREBRO, EL CEREBELO Y EL TRONCO CEREBRAL.
Que es y como funciona un ordenador
Un Ordenador es un Sistema conformado por programas y elementos electrónicos, que en su conjunto permiten procesar y ordenar información. Los programas se les conocen como Software y a los elementos electrónicos como Hardware.
Un ordenador es un sistema, formado por elementos eléctricos y programas, que permiten, ordenar y procesar la informacion
Composición del Cerebro
Cerebro
La corteza cerebral Está contenida en el cráneo gracias a numerosos pliegues y hendiduras. Solo un tercio de la corteza está expuesta superficialmente, el resto está oculto en la profundidad de los surcos. el cerebro se comuniquen más fácilmente, ya que están más cerca.
La corteza forma parte de lo que se conoce como sustancia gris, que es un componente del tejido cerebral que está mayoritariamente constituido por los cuerpos de las neuronas. Por debajo de la corteza encontramos la sustancia blanca, formada por prolongaciones nerviosas recubiertas de mielina que transmiten la información y permite que las neuronas de diferentes regiones del cerebro se conecten entre sí.
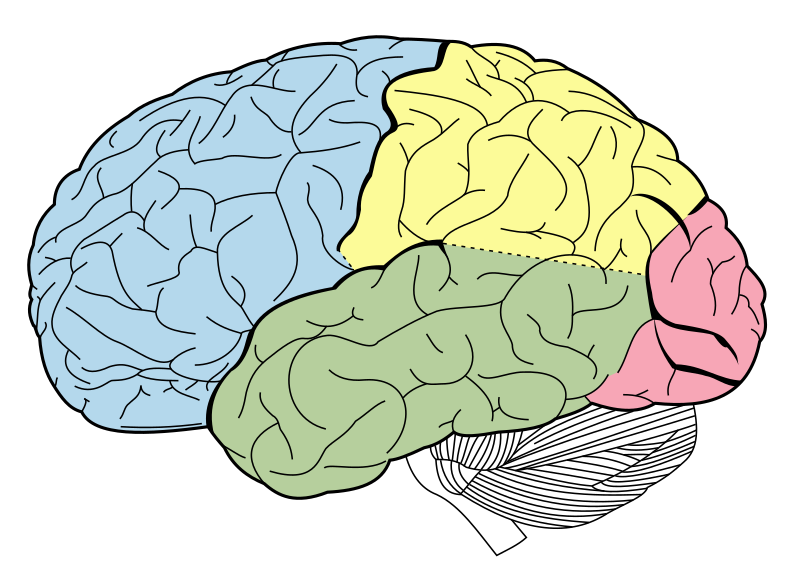
El cerebro está dividido en dos grandes partes, el hemisferio derecho y el izquierdo, que están conectados entre sí por un conjunto de fibras, que constituyen el cuerpo calloso. Cada uno de los hemisferios cuenta con cuatro lóbulos: frontal, parietal, temporal y occipital. Y cada lóbulo contribuye de manera diferente a las distintas funciones del cerebro. Por ejemplo, a grandes rasgos, el lóbulo occipital es fundamental para procesar la información visual, el parietal se encarga de integrar distintos tipos de información sensorial para guiar nuestros movimientos, el temporal nos ayuda a dar significado a la información sensorial, auditiva y visual, y permite muchos procesos relacionados con el lenguaje y, por último, el frontal actúa como un director de orquesta para planificar y ejecutar nuestros actos a partir de la información que recibe de diferentes regiones cerebrales y también participa en la producción del lenguaje. Además, en el lóbulo temporal también se encuentra el hipocampo, que tiene un papel crucial en el aprendizaje y en la memoria .
Además, en la parte más profunda de los hemisferios, encontramos diversos núcleos que, igual que la corteza, forman parte de la sustancia gris. Algunos de los más importantes son los ganglios basales y el tálamo, que, entre otras cosas, participan en funciones relacionadas con el procesamiento de información sensitiva y motora.
El cerebelo, ubicado en la parte postero inferior del cráneo, y mantiene el equilibrio y en la coordinación de los movimientos.
Tronco cerebral conecta el cerebro con la médula espinal y controla acciones corporales automáticas como el ritmo cardíaco, la tensión arterial y la respiración, así como el movimiento voluntario de los ojos, la lengua y los músculos de la cara, entre otros.
La médula espinal que comunica el cerebro con el resto del organismo, trasladando impulsos nerviosos desde el cerebro a distintas regiones del cuerpo.
La médula espinal produce una actuación por sí misma, sin que la información llegue a ser transmitida al cerebro. Es lo que conocemos como reflejos.
Las sinapsis comunican las. Son las responsables de la recepción de los inputs sensoriales provenientes del mundo exterior, así como de enviar órdenes a distintas partes del cuerpo y de transformar y transmitir las señales eléctricas que lo permiten.
Los componentes del cerebro son:
En el cuerpo celular está el núcleo donde se aloja el ADN y se forman las proteínas.
El axón transporta los mensajes electroquímicos.
Las dendritas o ramificaciones nerviosas son proyecciones cortas de la célula, como ramas, que establecen conexiones con otras células. Las dendritas reciben los mensajes a través de los neurotransmisores que liberan los axones de otras células nerviosas. En la parte inicial del axón de una neurona (donde se une al cuerpo neuronal) se genera un potencial de acción, un breve impulso eléctrico que viaja a lo largo del axón y provoca la liberación de neurotransmisores (son como mensajeros) en la sinapsis, el punto donde se produce esta liberación y la recepción del mensaje por otra neurona, permitiendo así la comunicación entre ellas.
La conexión entre las neuronas es fundamental para su propia subsistencia.
El tejido nervioso contiene otro tipo de células que forman la llamada neuroglia, cuya función consiste en dar mantener la homeostasis de las células nerviosas, aportarles los nutrientes necesarios y defender al tejido nervioso de virus u otros microorganismos.
Partes de un ordenador
Los ordenadores funcionan con transistores que reaccionan a impulsos de electricidad que pasan por sus circuitos, respondiendo siempre con el sistema binario,
Hardware:
Procesador: también conocido comúnmente como CPU (Unidad Central de Proceso), se encarga de llevar a cabo las instrucciones que contienen cada uno de los programas
Software:
Sistema Operativo: es un programa capaz de gestionar los recursos de los que dispone el ordenador. Es indispensable, ya que sin este ningún ordenador sería capaz de ejecutar función alguna.
Sistema Operativo:
Este se conforma por el núcleo del sistema (kenel), los controladores del dispositivo (drivers) y la interface de usuario (user interface).
El kenel se encarga de gestionar los recursos disponibles del ordenador, especialmente los del CPU y la RAM (Random Access, es la memoria principal y se usa para almacenar datos de acceso directo. También esta encardado de validar el acceso de los usuarios.
Los drivers (controladores del dispositivo) son generalmente programas que se encargan de definir como acceder a los dispositivos periféricos (que se encargan de las operaciones de entrada y salida que complementan lo que realiza el procesador, suelen dividirse entre Almacenamiento como disco duro, CD, etc., De Entrada como teclado, ratón, etc., De Salida como monitor e impresora o De Comunicación como tarjeta de red, tarjeta de Wireless etc.).
La user interface o interface de usuario es un conjunto de programas que permiten al usuario interactuar con el ordenador mayor comodidad, gracias a las herramientas necesarias que pone a su disposición.
Placa base del ordenador, donde se insertan sin cableado todos los componentes. Aventaja al cerebro, porque no tienen cablead, la placa base se encarga de la comunicación y conexión de sus elementos
Fisiologia del cerebro
Para prevenir su propia muerte, las neuronas, con la ayuda de la neuroglia, se tienen que mantener y remodelar constantemente.
La información desde nuestro cerebro hasta los músculos de
nuestras manos para contraer las o cómo se envía la información cuando nos pinchamos un dedo.
Las neuronas son células que se encuentran en el sistema nervioso y que se encargan de producir conducir y transmitir los impulsos nerviosos
Anatomia de las neuronas.
Lo mas llamativo son las dendritas que son ramificaciones con forma de espina que se encargan de recibir las señales o los impulsos nerviosos y enviarlos al soma que es el cuerpo de la
neurona el soma se continúa con una cola que se conoce o tiene el nombre de acción el axón va a continuar la transmisión del impulso nervioso y va a culminar en ramificaciones pequeñas que se van a unir a las dendritas de la próxima neurona .
El axón nos es la prolongación de ls neuronas que transmite el impulso nervioso. Está recubierto por ciertas células que se conocen como células de Schwann.
Estas células producen una sustancia que se conoce como mielina y que forma la vaina sobre el cilindroeje.
Es un aislante y cuya función es aumentar la velocidad con la
que se propaga el impulso nervioso
La mielina se rompe poeriodicamente y forma unos nódulos que se llamam, nodos de Ramier . Que actúan como transistores para potenciar el estimulo eléctrico o químico que conduce el cilindroeje.
No todas las neuronas presentarán vaina de mielina lo que determina que algunas neuronas envían los impulsos nerviosos más rápidos que otras
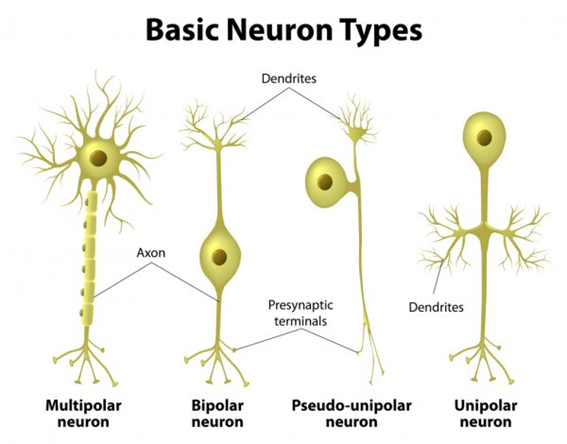
Según su forma las neuronas pueden dividirse en neuronas unipolares pseudo.unipolares bipolares y multipolar.
Las unipolares tienen un solo tipo de prolongación la pseudo unipolares tienen una prolongación la cual se divide en
una rama que entra al sistema nervioso central y otra que llega al sistema nervioso periférico, las bipolares presentan dos prolongaciones una que conduce los impulsos nerviosos hacia el
cuerpo celular de las neuronas y otra que aleja el impulso del cuerpo celular. Estas se encuentran en el epitelio olfatorio y las neuronas multipolares que se encuentran en el cerebro y en la médula espinal que presentan múltiples dendritas
Las neuronas también se pueden clasificar en neuronas aferentes que son las que comunican nuestro cerebro con el medio externo es decir las que nos envían las señales de dolor por ejemplo y también se conocen como neuronas sensitivas . Otras neuronas envían la información desde el cerebro hasta los músculos , se conocen como neuronas motoras y finalmente y producen el,movimiento
Las interneuronas son un conjunto de neuronas que se comunican entre sí
Que parecido tienen los computadores con el cerebro
Como funciona un ordenador
A pesar de que todos sabemos perfectamente como encender un ordenador, y este está en funcionamiento desde el momento en que presionamos el botón de encendido, Cuando lo encendemos, la corriente eléctrica llega a un transformador de fuerza o potencia. Este, a través del conector, distribuye los diferentes voltajes de trabajo a la placa base, incluyendo al CPU.
Inmediatamente después de que el CPU recibe corriente, envía una orden al chip de memoria ROM del BIOS (Basic Input/Output System – Sistema básico de entrada-salida), donde las rutinas del POST (Power-On Self-Test – Autocomprobación diagnóstica de encendido) o programa de arranque.
Si el BIOS (que contiene este conjunto de instrucciones grabadas en su memoria) no existiera, el sistema del ordenador no podría cargar la información y parte de los ficheros del Sistema Operativo en la memoria RAM, que se requieren para iniciar el arranque y permitir que se puedan utilizar los programas instalados.
Luego de que los BIOS recibe la orden del CPU, el POST lleva a cabo una secuencia de pruebas para diagnosticar si la tarjeta de video, la memoria RAM, las unidades de discos, el teclado, el ratón y otros dispositivos de Hardware que están conectados al ordenador, se encuentran en condiciones de funcionar correctamente.
Cuando el BIOS es incapaz de detectar un determinado dispositivo o directamente detecta fallas en alguno de estos, emitirá una serie de sonidos en forma de pitidos o “beeps”, haciendo aparecer en la pantalla del monitor mensajes de error, que indicaran que la existencia del problema.
En caso de que este no detecte ningún problema en medio de la revisión, se dirigirá hacia el sector de arranque del disco duro (boot sector) para proseguir con el arranque del ordenador.
Los errores encontrados por el BIOS, se clasifican en “grave” o “no grave”. En caso de que el error se clasifique como “no grave” solo se mostrara un mensaje de texto en la pantalla o los “beeps”, sin que el proceso de encendido del Sistema Operativo se vea afectado. En caso de que el error sea “grave”, el sistema se detiene y el ordenador se quedara bloqueado, en este caso, lo mejor es revisarlo y repararlo ya que, lo más probable es que algún dispositivo del Hardware no esté funcionando
Procesador de un ordenador
TRANSISTOR la invención de los transistores revolucionó la civilización humana como
ninguna otra tecnología en el corazón de un teléfono inteligente se encuentra un procesador y este procesador contiene alrededor de 2 mil millones de transistores que hacen estos
dispositivos increíblemente pequeños
Los transistores actuan como un interruptor pueden amplificar una señal débil la amplificación
Los transistores en la actualidad están hechos de silicio, pero el grafeno parace que tiende a sustiruirlo. Son semiconductores como el silicio cada átomo del silicio se enlaza con 4 átomos de silicio vecinos el silicio tiene 4 electrones en su capa de valencia reemplazamos este átomo de silicio con una figura sonriente de 4 manos cada mano tiene un electrón cada uno de estos electrones se comparte con un átomo de silicio vecino esto se conoce como enlace covalente actualmente los electrones están en su banda de valencia si el silicio puro tiene que conducir la electricidad los electrones tienen que absorber algo de energía y convertirse en electrones libres por lo tanto el silicio puro tendrá una baja conductividad eléctrica
Una pequeña corriente de base se amplifica a una alta corriente de colector
Quizás este escrito seria mas creíble si compararamos un ordenador con el cerebro del Australopitecus.
Pero a partir del Neandeltal y mas de Sapiens Sapiens, ya no tienen muchas similitudes.
Cuando el hombre convierte sus ideas en realidades, un fenómeno magnanimo ha aparecido que aun no encontramos su explicación.
Pero seguro que la tienen
La imágenes y el contenido de esta narración esta copiada de Internet.
La unión de lo orgánico con lo funcional, es entendible, pero enlazado con la vida psíquica invita a imaginar y esto nuestro cerebro no lo tiene aun orquestado
La evolución es interpretada de dos formas
La primera es referirnos a unas especies como antiguas, y más antiguas que otras. Aparecieron antes que las demás. Decimos, por ejemplo, que las esponjas son muy antiguas.
La segunda es que todos descendemos del mismo linaje hasta las primeras formas de vida que aparecieron sobre la faz de la Tierra y fueron capaces de dejar descendencia tras de sí generación tras generación y por tanto, somos herederos de aquellas formas , por lo tanto, todos los linajes, sean del reino que sean, o de la familia o género que sean, tienen la misma antigüedad, tanta como la vida terrestre tiene.
Esto quiere decir, que unos lo interpretan como una sucesión de formas biológicas sucesivas hasta llegar al homínido, y otros, como todo al mismo tiempo.
Un grupo de investigadores ha reconstruido la evolución del hipotálamo, una región del cerebro, hasta unos antepasados marinos similares al gusano. Esta labor, que ha sido financiada por la UE, ayuda a esclarecer la evolución del cerebro de los vertebrados.
Posiblemente esto nos llevaría al lenguaje romántico de muchos investigadores, que tienen una base de partida sólida y casi objetiva y el resto lo imaginan.
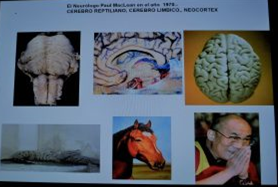
Quiero discutir como el cerebro Triuno de Mcclean , tiene una objetividad anatómica, aquí se expresan tres cerebros, pertenecientes a reptiles, mamíferos y homínidos, al mismo tiempo y sobre todo tienen una anatomía indiscutible, de estos tres grupos de seres viviente.
El cerebro del hombre esta compuesto por tres cerebros.
EL CEREBRO TRIUNO DE MACCLEAN Paul D. MacLean (1 de mayo de 1913 – 26 de diciembre de 2007) fue un médico norteamericano y neurocientífico que hizo importantes avances en los campos de la psicología y la psiquiatría : Su teoría evolutiva del cerebro triúnico propone que el cerebro humano es en realidad tres cerebros en uno: el reptiliano, el de los mamiferos y el del hombres. James Papez ya había acuñado esta teoría. El Neurólogo Paul MacLean fue el primero en proponer que el cerebro humano tiene tres porciones que son la suma de los cerebros que han pertenecido a otros animales en la evolución y cada una de ella creció encima de la otra. A lo largo de su evolución, el cerebro humano adquirió tres componentes que fueron surgiendo y superponiéndose. 1. Cerebro primitivo (arquipálio), constituido por la estructuras del tronco cerebral: Bulbo, cerebelo, puente y mesencéfalo, con el más antiguo núcleo en la base, el globo pálido y bulbos olfatorios. Se dice que corresponde al cerebro reptiliano, también llamado complejo-R por Paul MacLean. 2. Cerebro intermedio (paleopálio), formado por las estructuras del sistema límbico. Y se corresponde al cerebro de los mamíferos inferiores. 3. Cerebro superior o racional , el Neopálio, situado en la capa superior), que comprende la mayor parte de los dos hemisferios cerebrales (formado por el neocórtex) y algunos grupos neuronales subcorticales. Este último solo es compartido por los mamíferos superiores, incluyendo a los primates y el hombre. Los tres cerebros están interconectados como computadoras biológicas y cada uno tiene su propia inteligencia especial, su propia subjetividad, su propio sentido del tiempo y del espacio y su propia memoria
El Complejo Reptiliano El Complejo-R se compone del tronco cerebral y del cerebelo. Su objetivo está estrechamente relacionado con la supervivencia física real y el mantenimiento del cuerpo. Los tres cerebros se desarrollan superponiéndose durante la evolución embrionaria del feto. Y también cronológicamente en la evolución de las especies ,filogenia, desde el lagarto hasta el homo sapiens. En palabras de MacLean, son como tres computadoras biológicas que, aunque íntimamente interconectadas, conservan cada una sus propias formas peculiares de inteligencia, subjetividad, sentido del tiempo y del espacio, memoria, motricidad y otras funciones menos específicas. La parte más primitiva del cerebro básico, es el cerebro instintivo y reptiliano y esta formado por los ganglios basales, el tallo cerebral y el sistema reticular. Es esa parte la que se ocupa de las actividades intintivas. Se aloja en el tronco cerebral y se calcula que se desarrolló hace unos 500 millones de años. Se encuentra presente primordialmente en los reptiles, que son las especies animales con un menor desarrollo cerebral. El suyo, está diseñado para manejar la supervivencia desde un sistema binario: huir o pelear, con muy poco o ningún proceso sentimental. Tiene un papel muy importante en el control de la vida instintiva y se encarga de autorregular el organismo. Este cerebro no está capacitado para pensar, ni sentir. Su función es la de actuar, cuando el estado del organismo así lo demanda. La conducta animal e instintiva está en gran medida controlada por esta área del cerebro. Se trata de un tipo de conducta instintiva programada y poderosa y, por lo tanto, es muy resistente al cambio. Es el impulso por la supervivencia: comer, beber, mantener la temperatura corporal, sexo, territorialidad, necesidad de cobijo y de protección. Es un cerebro funcional, territorial, responsable de conservar la vida y el responsable de las mayores atrocidades. Nos sitúa en el presente, sin pasado ni futuro y por tanto es incapaz de aprender o prever. No piensa ni siente emociones y es pura impulsividad. En el cerebro reptiliano se procesan las experiencias primarias, no verbales, de aceptación o rechazo. Aquí se organizan y procesan las funciones que tienen que ver con el hacer y el actuar. Es el responsable de las conductas automáticas, tales como las que se refieren a la preservación de la especie y a los cambios fisiológicos necesarios para la sobrevivencia. El sistema básico o reptiliano controla la respiración, el ritmo cardíaco, la presión sanguínea e incluso colabora en la continua expansión-contracción de nuestros músculos. Este primer cerebro es sobre todo como un guardián de la vida, pues en él están los mayores sentidos de supervivencia y lucha. Y además, mantiene la interrelación con los poros de la piel, los cuales son como una especie de interfase que poseemos con el mundo externo. Este primer cerebro es nuestro agente avisador de peligros para todo el cuerpo. Permite la adaptación con rapidez por medio de respuestas elementales poco complicadas emocional o intelectualmente. Esta conducta no está basada en consideraciones basadas en las experiencias previas ni en los efectos a medio o largo plazo. Las conductas de las personas calificadas como de psicópatas, las que carecen de sentimientos de culpa y de paranoicos se ajustan a este patrón de conducta. En la psicopatía se juega el papel de depredador y en la paranoia el de presa. Es en este primer cerebro donde las adicciones son muy poderosas, tanto a algo como a alguien o a una forma de actuar. Por decirlo de alguna forma rápida, este primer cerebro es una herencia de los períodos cavernarios, donde la supervivencia era lo esencial.
Desempeña un papel crucial en el establecimiento de territorio, la reproducción y la dominación social. Las características primordiales de los comportamientos del Complejo-R es que son automáticos, tienen una cualidad ritual, y son muy resistentes al cambio.
SISTEMA LÍMBICO La parte media del cerebro es llamada “sistema limbico ” Puede también ser llamado el paleopallium o el cerebro intermedio o cerebro de los viejos mamíferos. Aquí se asientan las emociones y los instintos, alimentación, lucha y huida, y comportamiento sexual. En este sistema se acumula lo agradable o desagradable y la supervivencia depende de evitar el dolor y obtener el placer.
El sistema Limbico en su totalidad parece ser el asiento primario de la emoción, de la atención, y de las memorias afectivas. Anatómicamente incluye el hipotálamo, el hipocampo, la amigdala.
Los Budistas afirman que aquí se alojan la determinación de la valencia positiva o negativa hacia algo y el comportamiento creativo. Las conexiones de este cerebro con el neocórtex son amplias en ambos sentidos de forma que las reacciones son una mezcla en sus respuestas de lóbulo limbico y telencefalo Según MacLean el sistema Limbico tiene una tendencia dogmática y paranoica y la base biológica para la tendencia del pensamiento como sensación subordinada a racionalizar deseos. En opinión de McClean este cerebro intermedio aloja juicios de valor en vez de alojarse en el neocortex, motivando o produciendo con frecuencia confusiones . EL NEOCORTEX Neocortex es la corteza del cerebro también conocido como el cerebro neo mamífero ,neo mamalian, y aloja lo racional y superior y se extiende a prácticamente a los hemisferios cerebrales y algunos grupos neuronales subcorticales.
Es la ultima adquisición de los homínidos y ocupa dos tercios de la masa total del cerebro. Todos los animales también tienen un neocortex, es relativamente pequeño, con escasos pliegues y menor complejidad y desarrollo, de forma que anatómicamente los tres modelos no estan perfilados de forma que a nivel de anécdota, se puede explicar como los mamíferos pueden aprender aunque con dificultad. El cerebro de los primates y, por lo tanto, de la especie humana, aloja las funciones cognoscitivas más altas que distinguen a hombre de los animales. MacLean llama a la corteza del cerebro “la madre de la invención y el padre del pensamiento abstracto “. La corteza se divide en los hemisferios izquierdos y derechos. La mitad izquierda de la corteza controla la parte derecha del cuerpo y el cerebro derecho, el lado izquierdo del cuerpo. También, el cerebro derecho es más espacial, abstracto, musical y artístico, mientras que el cerebro izquierdo más linear, racional, y verbal.
Hoy se cree: que en el funcionamiento del cerebro no importa tanto la función que realizan las partes del cerebro por sí solas como el modo en el que se conectan entre sí para trabajar en conjunto y en tiempo real. Además, por lo que se sabe la evolución no va haciendo que componentes nuevos vayan integrándose sobre los antiguos, sin alterarlos.
Cada vez que aparece una mutación hace que un rasgo se generalice, altera el funcionamiento del organismo en su totalidad y el modo en el que funcionan las partes que habían evolucionado antes, no se limita a “expandir” capacidades. Es por eso que la idea de que órganos cerebrales “encargados de lo racional” se acoplan sobre los anteriores no ha sido bien aceptada. Las funciones que supuestamente realizaban cada uno de los tres cerebros definen bien el comportamiento característico de los grupos de animales que, según él, representan el momento de la evolución en el que aparecieron estas estructuras. Posiblemente, cada uno de estos cerebros tiene funciones del cerebro anterior y del posterior en forma de excitación o inhibición de los cerebro que están encima o bajo el anterior y la alteración de cada uno libera al inferior como estableció Sherrington. La visión actual de los ganglios basales (que formarían parte del cerebro reptiliano) es que no se activan por acciones programadas genéticamente, sino que están asociados a la realización reiterada de movimientos voluntarios que después de haber sido muy practicados, se han vuelto automáticos, como el tan cacareado ejemplo de ir en bicicleta. Los seres humanos, hemos evolucionado desde siempre, para adaptarnos a nuestro entorno. La división de nuestra mente en 3 partes, parece ser la teoría más aceptada, por su forma de aplicarse en la vida real. Nuestra mente es nuestra mejor ventaja evolutiva. Lo cierto es que anatómicamente coexisten estos tres cerebros en el homo, y funcionalmente sucede esto.
El HIPOTALAMO es la glándula productora de hormonas que regula toda nuestra biología y es necesario hacer un esfuerzo para entender que esta glándula sea capaz de fabricar hormonas selectivas para cada función organica, pero como siempre tropezamos con las funciones superiores.
Como una hormona es capaz de sublimar nuestra biología ¿ que es reproducir la frase de Francisco Mora de una manera dramática dice “” como un montón de neuronas enmarañadas unas con otras pueden dar lugar a un a un individuo que piensa y siente, que llora y ríe y con ello levanta su mirada hacia el infinito universo y se pregunta por su existencia y su sentido
HIPOTALAMO
El hipotálamo de los vertebrados produce hormonas, señales químicas que controlan el crecimiento, el metabolismo, la reproducción y muchos otros procesos fisiológicos. Los insectos y los gusanos nematodos también producen hormonas, pero el aspecto de éstas es muy distinto al de las hormonas de los vertebrados, lo que hacía suponer a la comunidad científica que estas regiones del cerebro secretoras de hormonas habían aparecido con posterioridad a la separación evolutiva de los vertebrados y los invertebrados.
Sin embargo, la comunidad investigadora descubrió después hormonas similares a las de los vertebrados en gusanos y moluscos, lo que indica que estas estructuras podrían ser más antiguas de lo que se pensaba.
En un artículo publicado en la revista Cell, científicos del Laboratorio Europeo de Biología Molecular (EMBL) y de la Universidad Libre de Berlín describen de qué modo compararon las células neurales secretoras de hormonas del pez cebra (un vertebrado) y del gusano anélido.
Hallaron similitudes asombrosas entre ambos grupos; ambas clases de células tenían un aspecto similar y se hallaban en la misma posición en los cerebros en desarrollo de las dos especies. Además, mostraban la misma configuración molecular. Estas similitudes no pueden atribuirse simplemente a una coincidencia y delatan un origen evolutivo común de estas células.
«Es probable que ya existieran en los últimos antepasados comunes conocidos de los vertebrados, los insectos y los gusanos», Las células estudiadas son multifuncionales; además de ser capaces de secretar hormonas, tienen propiedades sensoriales, ya que responden a la luz y a ciertas sustancias químicas. Los investigadores creen que estos tipos de células «sensoriales-neurosecretoras» están entre los tipos más antiguos de células nerviosas. Habrían permitido responder directamente a cambios en el entorno marino. Con el tiempo estas células multifuncionales fueron formando grupos a modo de centros cerebrales y se diversificaron en diversas diferentes especialidades, como se observa en el cerebro de los vertebrados modernos.
«Estos hallazgos cambian drásticamente el modo en que entendemos el cerebro», según Kristin Tessmar-Raible, autora principal del artículo. «Hasta ahora siempre lo veíamos como una unidad de procesado, similar a un ordenador que integra e interpreta la información sensorial que recibe. Ahora sabemos que el cerebro es en sí mismo un órgano sensorial y que es así desde tiempo inmemorial.
Lo único que aporta esto es que los limites de los parénquimas, en este caso el cerebro triuno, no es funcional, sino que se combinan las capacidades de cada uno y dan unas funciones superiores, independientemente de cuando han aparecido cada uno de los cerebro. Están junto, colaboran y la química no seria muy difícil de explicar
El problema viene cuando nos preguntamos. Que función superior las organiza y como hace esto?.
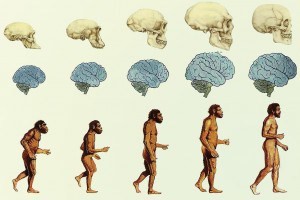
La hominización hace referencia a las diferentes transformaciones que el ancestro común de los primates y los homínidos sufrió a través de distintas especies hasta producir a los seres humanos como los conocemos en la actualidad el homo sapiens sapiens diferentes autores concuerdan entonces en que los cambios más resaltantes que ocurrieron en dicho linaje pueden resumirse en cinco puntos fundamentales el desarrollo del bipedalismo caminar sobre los dos miembros posteriores, el perfeccionamiento de la manipulación manual y la fabricación, el uso de herramientas, la modificación de la mandíbula y de los dientes, el aumento en la masa cerebral y los cambios en las cuerdas vocales, el desarrollo del lenguaje y de la comunicación.
El proceso de hominización comenzó en Africa hace más o menos 6 millones de años cuando algunos primates ancestrales dejaron rastros fósiles de su capacidad para el uso de cierto tipo de herramientas, las fases del proceso de hominización corresponden a las distintas especies de las que se guardan registros fósiles estas son las especies del proceso de hominización
Australopitecus
Estos son tal vez los hominoides más antiguos que se han encontrado a partir de los cuales se
piensa que surgieron los homínidos los australopitecos habitaban las sabanas africanas hace unos 4 millones de años y como se comentó caminaban erguidos sobre sus extremidades posteriores eran bípedos de acuerdo con los estudios paleontológicos de los fósiles, estos primeros hominoides tenían un aspecto y una contextura física más parecida a la de un simio que a la de un humano y de acuerdo con el tamaño de sus cerebros se cree que eran tan inteligentes como los chimpancés modernos.
De este género se conocen diversas especies que pudieron haber existido en el espacio temporal correspondiente a 2.5 millones de años atrás
El primer representante del género homo, es el homo habilis que existió hace 2.5 millones de años ya se comentó acerca de su capacidad de fabricar herramientas y de sus comportamientos sociales además de su mayor capacidad cerebral respecto a los australopitecos homo Ergaster.
El homo Ergaster vivió hace 1.8 millones de años y al igual que el homo habilis tenía la capacidad de fabricar herramientas con distintos propósitos entre ellos la casa lo que le proporcionó mas consumo de carne. Esta especie era probablemente, mayor que sus predecesores. De esta especie se han registrado fósiles tanto en Asia como en Europa y en el norte de Africa y se ha determinado que su capacidad craneal era aún mayor que la del homo Habilis .
Erectus al igual que el homo Ergaster y el homo Erectus vivió hace 1.6 millones de años y mantenía la habilidad para hacer herramientas y utensilios de cazadores . Estos homínidos lograron dominar el fuego y quizás se comunicaban con una suerte de lenguaje primitivo.
El homo Antecessor es considerado la especie más antigua del homínido hallada en Europa la cual habitó la tierra hace más o menos 900 mil o un millón de años, tenían cerebros más pequeños que el de los humanos modernos eran de contextura más grande que la de los homínidos anteriores y se cree que precedieron a los homo sapiens
Homo sapiens
Los representantes del homo sapiens fueron hallados entre Europa y Asia hace poco más de 200.000 años por lo que se piensa que convivieron en otros representantes del género tenían mayor capacidad craneal más de 1000 cc y podían fabricar herramientas y armas más elaboradas o sofisticadas. Construían sus viviendas, tenían cierta organización y practicaban rituales funerarios para sus difuntos
El Homo Sapiens Sapiens (el hombre que sabe que sabe), corresponde a los hombres modernos. Apareció hace 160.000 años como el hombre de Cromagnon que tenía una capacidad craneal de casi 2000 cc, los primeros registros fósiles sugieren una gran capacidad para elaborar herramientas y utensilios domésticos de piedra madera y hueso fueron los primeros en presentar complejas organizaciones sociales tribus y en aumentar el lenguaje y facilitar comunicación.
En esta especie surgió la creatividad y con ello el arte lo que se deduce por las pinturas rupestres halladas en las cuevas de Altamira en España’ posteriormente se dieron los procesos de culturización y de civilización los cuales marcaron un antes y un después en la historia de la humanidad tal como la conocemos actualmente
Las características de la hominización estuvieron definidas por algunos eventos característicos que ocurrieron en las distintas especies del linaje y que conllevaron al desarrollo del hombre moderno:
Estas características se resumen en:
La capacidad de caminar erguido sobre los miembros inferiores es tal vez una de las características más antiguas compartidas por todos los homínidos y que apareció en los ancestros más primitivos pertenecientes al género. Asimismo la angulación dde la base del cráneo, permitió alojar un cerebro mas grande.
Australopithecus, los registros fósiles los sitúan hace unos 4 millones de años que corresponden a una hembra del género descubierta en Etiopía en 1974 a quien bautizaron, como Lucy.
Los fósiles han permitido encontrar subtipos
Australopithecus africanus .
Australopithecus afarensis
Australopithecus ramidus y
Australopithecus annamenzies
También del mismo género la capacidad de caminar erecto implicó una serie de modificaciones importantes del esqueleto las cuales fueron necesarias para la transición de una vida sobre los árboles a una vida en las sabanas africanas .
Al mismo tiempo apareció el:
El alargamiento de los miembros inferiores y aplanamiento de las plantas de los pies el acortamiento de los miembros superiores incluyendo los dedos de las manos, el ensanchamiento de las palmas de las manos y desarrollo de un pulgar oponible, la reestructuración de la columna en forma de S para soportar la cabeza en posición vertical, el estrechamiento y fortalecimiento de la pelvis para soportar las vísceras y órganos internos.
En esta fase del proceso de hominización es bueno establecer que los hominoides pertenecientes al género australopithecus tenían cerebros pequeños rostros prominentes al igual que sus dientes y brazos mucho más cortos que las piernas y la anatomía de las manos que les permitió utilizar la capacidad de las manos con cierta precisión diferente a la de los simios y más similar a la de los miembros del género homo se observó por primera vez en Australopithecusy en consecuencia el uso de herramientas se incremento y perfeciono.
Afarensis una especie de hominoide que tenía manos de proporciones similares a las de los humanos pero con dedos mas gruesos que les permitía una mayor capacidad de agarre, que era superior a los simios.
La siguiente fase del proceso de hominización está representada por hallazgos paleontológicos de la especie homo Habilis.
Hay evidencia indiscutible de que la fabricación de herramientas de piedra ocurrió hace más de 2 millones de años, antes de la aparición de cerebros más grandes y complejos.
El homo Habilis es la primera especie del género homo que se conoce, eran homínidos de baja estatura menos de 1.50 metros con cerebros apenas más grandes y dientes más pequeños que los de los australopithecus. Se les llamo Habilis por su habilidad para tallar piedras
Después de habilis hace unos 1.8 millones de años y hasta hace 200 mil años vivieron sobre la tierra
El homo Erectus que construia herramientas más sofisticadas y que también aprendió a controlar el fuego, lo que tuvo gran significancia ecológica y les dio la posibilidad de cocinar sus alimentos , mantenerse caliente durante la noche en temporadas frías, ahuyentar depredadores e iluminar sus caminos en la oscuridad.
Tenía una capacidad cerebral aún mayor que la del homo habilis y con ellos aparecieron las primeras
organizaciones sociales que permitieron un gran desarrollo de la comunicación a través del lenguaje hablado en los y dispersarse por zonas más templadas de Eurasia.
Existe evidencia de que los neandertales pertenecientes a la especie homo sapiens neanderthalensis que habitaron Europa y Asia occidental hace unos 6000 años, tenían prácticas religiosas.
Los Cromañones, los representantes europeos de homo sapiens sapiens, construían sus viviendas y mantenían comunidades estables hace unos 40 mil años.
Domesticaron animales y plantas y aparecio la agricultura y las primeras civilizaciones no Tardarpn en aparecer.
Modificación las mandíbulas y de los dientes, las diferencias entre las mandíbulas y los dientes de los simios y los homínidos son bastante importantes.
Los primeros tienen dientes esmaltados, gruesos y grandes con caninos y molares prominentes así como una gran mandíbula y sus músculos asociados en comparación con los australopithecus, por ejemplo los simios tienen una mayor relación entre el área comprendida por los dientes y el peso corporal australopithecus Radimus tenía dientes
parecidos a los de algunos representantes del género como dientes pequeños con una delgada capa de esmalte y caninos cortos similares a los chimpancés. El australopitecus Afarensis tenía rasgos compartidos entre simios y homínidos Incisivos como los de los chimpancés pero caninos como
los de los australopithecus africanus. Homo habilis tenía dientes más o menos parecidos a los de australopithecus Afarensis con una una reducción gradual del tamaño de los dientes frontales y un incremento del tamaño de los dientes laterales más cercanos a las mejillas, se piensa que esto estaba relacionado con ciertos cambios climáticos los
cuales pudieron haber modificado la composición vegetal y animal del entorno en el que estos organismos habitaban modificando también sus hábitos alimenticios aumento de la masa cerebral .
La evolución del cerebro desde los primeros homínidos hasta el hombre moderno y la angulación de la base del craneo, ocurrieron poco después de la aparición del bipedalismo y una vez los cambios en la dentadura y las mandíbulas culminaron con el aumento de la masa cerebral
También se dio un aumento del tamaño corporal y este proceso de encefalización también estuvo acompañado con otros cambios característicos de los homínidos homo Habilis tenía un volumen cerebral mucho mayor que el de los Australopithecus pero distintos estudios computarizados han comprobado que el incremento no fue gradual el periodo
entre hace 4 y 2 millones de años mostró cambios insignificantes del volumen cerebral al menos en lo que respecta a Australopithecus Afarensis y Australopithecus Africanus , compartían cerebros de menos de 450 cc, mientras que los Homo Habilis hace 2 o 1.5 millones de años, tenían cerebros entre 650 y 700 cc.
Se deduce que la evolución del cerebro de los homínidos ocurrió en realidad en un momento de la historia hace unos 2 o 1.5 millones de años.
La comunicación mantenida por el aparato fonético de los humanos, tiene dos componentes, el sistema subtotal bajo de la glotis que consiste en los pulmones y sus músculos y la laringe que comunica el sistema subletal con el tracto superior de esta. Las vías supra laríngeas humanas son diferentes a las de otros primates pues en los humanos el paladar está hacia atrás y la laringe está hacia abajo lo que permite la construcción única de una vía supra laringe a diferente a la de otros primates más aún el hecho de que la lengua humana pueda moverse en el espacio definido por el paladar y la columna vertebral permite generar los patrones de frecuencia que consiguen los sonidos de las vocales y las consonantes
Las estructuras y el mecanismo de control nervioso necesario para el establecimiento del habla humana evolucionaron hace 1.8 millones de años en homo Erectus de acuerdo con lo que sugieren la anatomía comparada entre los primates actuales y los fósiles del oso.
El lenguaje es una adaptación única de los humanos pues evolucionó en el cerebro humano aunque no existen evidencias anatómicas de la adquisición de ningún órgano nuevo diferente al de los predecesores del hombre moderno hecho que dificulta el estudio de su origen. Los antropólogos difieren respecto a cuando apareció por primera vez el lenguaje, algunos afirman que su origen acompañó a la aparición del homo sapiens moderno con el tamaño del cerebro moderno y el tracto vocal descendente completamente formado, otros por el contrario ubican la aparición del lenguaje en los tiempos de homo habilis con los primeros registros de las herramientas y el comienzo del aumento en la masa cerebral sin embargo las diferentes adaptaciones encontradas para el desarrollo del lenguaje y su profunda integración con la naturaleza humana parecen indicar que este tuvo su origen hace muchos millones de años y ha sido sugerido que las primeras formas e incluían cantos y gestos.
Y aquí estamos nosotros, que nos distinguimos, porque ideamos y desde esta ideas construimos materias y dispositivos sociales complejos, que a su vez nos ayudaron a dar mas inteligencia a nuestra creatividad.
Estamos aquí en espera de nuevos acontecimientos. El ejercicio de las maquinas y sobre todo de la inteligencia artificial, no cabe duda que dará algo nuevo y esperamos que esté cargado este algo, de bondad y amor a los demás y a todo en general
Describir la anatomía del cerebro de forma fragmentada puede ser un error . sin embargo dada la evolución de la cirugía focalizada es imprescindible el conocimiento detallado de algunas vías , lo que proporciona conocimiento imperfecto, pero util .
Intento describir las proyecciones de la médula hasta el tálamo y a la corteza cerebral
El tracto espinotalámico o vía espinotalámica, sistema anterolateral o el sistema ventrolateral) es una vía sensorial desde la piel hasta el tálamo. Desde el núcleo ventral posterolateral en el tálamo, la información sensorial se transmite hacia arriba a la corteza somatosensorial de la circunvolución poscentral.
El tracto espinotalámico consta de dos vías adyacentes: anterior y lateral. El tracto espinotalámico anterior transporta información sobre el tacto crudo. El tracto espinotalámico lateral transmite dolor y temperatura.
En la médula espinal, el tracto espinotalámico tiene una organización somatotópica. Esta es la organización segmentaria de sus componentes cervical, torácica, lumbar y sacra, que está dispuesta desde la más medial hasta la más lateral, respectivamente.
Hay dos partes principales del tracto espinotalámico (STT):
El tracto espinotalámico lateral transmite dolor y temperatura.
El tracto espinotalámico anterior (o el tracto espinotalámico ventral) transmite un tacto crudo y una presión firme.
El tracto espinotalámico, al igual que el tracto dorsal de la columna medial lemniscus, utiliza tres neuronas para transportar información sensorial desde la periferia hasta el nivel consciente en la corteza cerebral.
Las neuronas pseudounipolares en el ganglio de la raíz dorsal tienen axones que conducen desde la piel a la médula espinal dorsal donde ascienden o descienden uno o dos niveles vertebrales a través del tracto de Lissauer y luego hacen sinapsis con neuronas secundarias en la sustancia gelatinosa de Rolando o el núcleo propio. Estas neuronas secundarias se llaman células del tracto.
Los axones de las células del tracto cruzan (decusado) hacia el otro lado de la médula espinal a través de la comisura blanca anterior, y hacia la esquina anterolateral de la médula espinal (de ahí que el tracto espinotalámico sea parte del sistema anterolateral). La decusación generalmente ocurre 1-2 segmentos del nervio espinal por encima del punto de entrada. Los axones viajan a lo largo de la médula espinal hasta el tallo cerebral, específicamente la médula ventromedial rostral.
Los tipos de información sensorial transmitida a través del tracto espinotalámico se describen como sensación afectiva. Esto significa que la sensación va acompañada de una compulsión a actuar. Por ejemplo, un picor se acompaña de la necesidad de rascarse, y un estímulo doloroso nos hace querer alejarnos del dolor. [Citación necesitada]
Hay dos subsistemas identificados:
En el sistema nervioso, el sistema anterolateral es una vía ascendente que transmite dolor,1 temperatura (sensación protopática) y tacto crudo desde la periferia al cerebro. Comprende tres vías principales:
Nombre
Destino
Función
Tracto espinotalámico (lateral y anterior)
Tálamo
Importante en la localización de estímulos dolorosos o térmicos
A diferencia de los axones de las neuronas de segundo orden en la vía posterior del lemnisco de la columna medial, los axones de las neuronas de segundo orden en los tractos espinotalámicos se cruzan a cada nivel segmentario en la médula espinal. Este hecho ayuda a determinar si una lesión está en el cerebro o en la médula espinal. Con lesiones en el tallo cerebral o más altas, las deficiencias en la percepción del dolor, la sensación táctil y la propiocepción son todas contralaterales a la lesión. Con las lesiones de la médula espinal, sin embargo, el déficit en la percepción del dolor es contralateral a la lesión, mientras que los otros déficits son ipsilaterales. Ver el síndrome de Brown-Séquard.
Las lesiones unilaterales generalmente causan anestesia contralateral (pérdida de dolor y temperatura). La anestesia normalmente comenzará 1-2 segmentos por debajo del nivel de la lesión, debido a las fibras sensoriales que se transportan por el tracto dorsal-lateral de Lissauer hasta varios niveles al ingresar a la médula espinal, y afectará todas las áreas del cuerpo caudal. Esto se prueba clínicamente mediante el uso de pinchazos.
PROYECCIONES TÁLAMO-CORTICALES
Son la vía de entrada a la corteza cerebral de información sensorial procedente de la periferia. Además, constituyen un relevo de información entre áreas corticales estableciendo un control del flujo de información en la comunicación cortico-tálamo-cortical. Lejos de ser similares, las neuronas talámicas, agrupadas en diferentes núcleos, poseen una diversidad en cuanto a sus arquitecturas axónicas conectivas, lo que, en último término, determina su función. En la corteza somatosensorial primaria de roedores, las sinapsis ‘lemniscales’ o ‘específicas’ procedentes de la porción dorsomedial del núcleo ventral posteromedial talámico (VPMdm) se establecen con las neuronas de capa 4, mientras que el núcleo posterior (Po), ‘paralemniscal’ o ‘multiespecífico’, conecta con las neuronas piramidales de capa 5a. A pesar del reducido número de botones tálamo-corticales, su sinapsis es altamente eficaz, lo cual podría depender de las características estructurales que presentan dichos botones. A través de un abordaje de microscopía electrónica, se han reconstruido y analizado cuantitativamente axones anterógradamente marcados, de ambos núcleos talámicos, en sus correspondientes capas corticales. La mayoría de las sinapsis se han localizado en botones, los cuales han mostrado diferencias en cuanto a su tamaño, número y tamaño mitocondrial y número de vesículas sinápticas. Además, los botones han establecido contacto sináptico en un 80% con espinas dendríticas, algunas de las cuales forman grandes protrusiones dendríticas incluidas por la membrana del botón presináptico. Se ha observado una densidad postsináptica de tamaño similar en ambas poblaciones, sin embargo, existe una mayor porcentaje de botones multisinápticos en axones procedentes de VPMdm. En conjunto, las características estructurales de los elementos pre- y postsináptico contribuyen al diferente fenotipo funcional observado para los axones de VPMdm como núcleo talámico de ‘primer orden’, y para los axones de Po como núcleo talámico de ‘orden superior’ implicado en la modulación de la transmisión sináptica. Palabras clave: Tálamo, Corteza somatosensorial, Diversidad sináptica, Microscopía electrónica de transmisión, Espinas dendríticas.
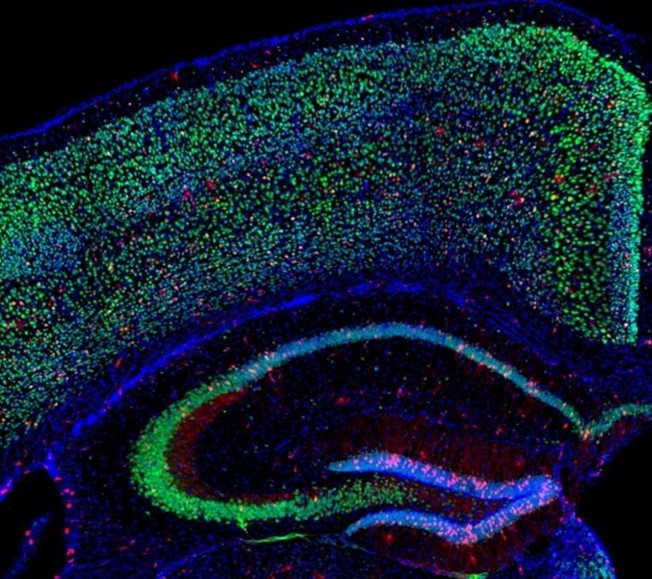
EL DÉFICIT DE OXÍGENO ESTIMULA LA FORMACIÓN DE NUEVAS NEURONAS
Hipoxia funcional en el cerebro: imagen que muestra la corteza y el hipocampo de un ratón con hipoxia inducida (déficit de oxígeno). Pueden apreciarse las neuronas hipóxicas en verde y con puntos rojos. Crédito: Instituto Max Planck de Medicina Experimental.
El déficit de oxígeno mientras se realiza actividad física o mental puede ser altamente perjudicial para todo el cerebro, desembocando en daños neuronales irreparables. Sin embargo, un nuevo estudio ha descubierto que una disminución controlada del oxígeno genera efectos positivos, estimulando el desarrollo de nuevas células cerebrales y favoreciendo sus conexiones o sinapsis.
Un grupo internacional de investigadores liderado por especialistas del Instituto Max Planck de Medicina Experimental en Gotinga, Alemania, ha demostrado en ratones que la ACTIVIDAD FÍSICA Y MENTAL EXIGENTE DESENCADENA UNA “HIPOXIA FUNCIONAL” O DÉFICIT DE OXIGENO, QUE SI ES CONTROLADA EFICAZMENTE PODRÍA TENER CONSECUENCIAS POSITIVAS EN TODO EL CEREBRO.
Según una nota de prensa, la hipoxia controlada logra estimular el crecimiento de nuevas sinapsis y neuronas, tanto en regiones específicas del cerebro, como el hipocampo, asi como en la totalidad de la estructura cerebral. Los científicos creen que este fenómeno podría explicar en parte los beneficios que conlleva la actividad física y mental durante la vejez, retrasando los efectos del envejecimiento en el cerebro.
Hipoxia controlada
En la investigación, publicada recientemente en la revista Molecular Psychiatry, los científicos sometieron a diferentes exigencias a un grupo de ratones modificados genéticamente. Los roedores debían concentrarse para correr sobre ruedas especialmente preparadas. Cuando se producía un déficit de oxígeno, un tinte fluorescente se activaba a partir de una proteína introducida, haciendo visible los cambios en las imágenes cerebrales.
Al comparar los resultados obtenidos en el grupo de estudio con otros ratones que no desarrollaron actividad física y mental, los científicos comprobaron que los roedores con hipoxia inducida mediante el ejercicio mostraban una activación del factor de crecimiento eritropoyetina (Epo) en el cerebro.
Dicho proceso se conoce por su efecto estimulante sobre los glóbulos rojos, pero al mismo tiempo promueve la formación de células nerviosas y su interconexión en el cerebro en forma de sinapsis. Además, los especialistas constataron que las neuronas eran especialmente beneficiadas por el efecto estimulante, mucho más que sus células auxiliares o gliales.
Cambios locales e integrales
La hipoxia controlada que se genera a partir de un desafío cognitivo o motor produce cambios en todo el cerebro, pero las neuronas “hipóxicas” se desarrollaron en mayor medida en el hipocampo. Más allá de esta ventaja localizada, los resultados muestran un modelo integral de neuroplasticidad, según los especialistas.
Esto quiere decir que una actividad neuronal asociada a una tarea específica desencadena una hipoxia leve como respuesta local concreta en diferentes regiones cerebrales, así como un efecto integral en todo el cerebro, que comprende neuronas y células no neuronales.
Aplicaciones
Teniendo en cuenta que la actividad física y mental promueve distintos procesos que logran mejorar la función cerebral global, incluyendo el estado de ánimo y el bienestar emocional, el hallazgo de este grupo de neurocientíficos podría desembocar en nuevas instancias terapéuticas a partir de los beneficios de la hipoxia controlada.
Ahora, los expertos trabajan en la adaptación del modelo desarrollado en roedores para que puede ser probado en seres humanos. Piensan aplicarlo a un grupo de estudio enfocado en una actividad concreta: los ejercicios con bicicletas estáticas.
Si se logran comprobar los mismos cambios en el ser humano, el nuevo enfoque podría ser especialmente importante para el tratamiento de enfermedades neurodegenerativas, pero al mismo tiempo podría ayudar a retrasar los efectos del envejecimiento en el cerebro en personas de edad avanzada e, incluso, funcionar como un potenciador de la actividad cerebral en personas jóvenes y sanas.
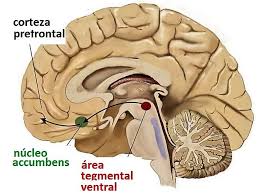
El núcleo Acumbens esta de moda, ha pasado de no ser conocido a serlo y mucho
¿Dónde está el núcleo accumbens?
El núcleo accumbens es una estructura cerebral subcortical, situada en el punto en que núcleo caudado y putamen se juntan con el septum. Este núcleo forma parte de la zona ventral del cuerpo estriado, siendo uno de los núcleos que configuran los ganglios basales.
.
Partes de esta estructura
El núcleo accumbens se ha dividido tradicionalmente en dos secciones, zona central y corteza, debido a sus diferentes conexiones con otras áreas cerebrales y a su mayor vinculación a lo emocional o a lo motor.
1. Corteza (Shell)
Esta parte del núcleo accumbens se caracteriza por su elevado número de conexiones con el sistema límbico y el hipocampo, recibiendo tanto dopamina como serotonina y glutamato de diversas áreas cerebrales.
Se trata pues de la parte más vinculada a las emociones de esta estructura. También tiene muchas conexiones provenientes del frontal, enviando el núcleo accumbens la información recogida al tálamo y recibiendo de vuelta a la zona central del núcleo accumbens.
2. Zona central (Core)
La zona central del núcleo accumbens tiene funciones principalmente vinculadas a la motricidad, estando conectado a los ganglios basales, la sustancia negra y la corteza motora.
Funciones principales
La localización de esta estructura y las conexiones que mantiene con diferentes áreas cerebrales hace que el núcleo accumbens sea una estructura de gran importancia.
Si bien muchos de ellos son compartidos por el resto de ganglios basales, algunos de dichos procesos en los que el núcleo accumbens tiene especial participación son los siguientes.
Funciones en la que interviene.
1. Integración emoción-motivación-acción Se lo considera involucrado principalmente en el sistema de recompensa o de refuerzo conductual positivo.
Cuando se estimulan eléctricamente cierto grupo de neuronas de esta área o cuando se presentan los llamados reforzadores conductuales positivos naturales (RPN), tales como la presencia de alimentos agradable dinero, sexo, etc., se produce la liberación de dopamina y con ello la activación de receptores dopaminérgicos en este núcleo.
La respuesta de recompensa no sólo se produce por la estimulación cerebral producida por los RPN, sino por la exposición repetida a drogas (cocaína, anfetaminas, heroína, alcohol, nicotina), siendo uno de los mecanismos involucrados en la adicción a las mismas. Es decir, las drogas adictivas pueden activar los mismos grupos neuronales que los RPN.
Las interconexiones del accumbens con la amígdala cerebral sugieren un alto contenido emocional en la toma de decisiones que implican recompensa.
De igual manera, sus abundantes interconexiones con las áreas de asociación frontal y prefrontal indican un papel importante en la planificación de comportamientos complejos como la expresión de la personalidad, los procesos de toma de decisiones y en la ejecución de un comportamiento social adecuado al contexto que se vive en el momento.
Se activa selectivamente durante la percepción de imágenes agradables, excitantes emocionalmente y durante las imágenes mentales de agradables escenas emocionales.
Es una interface cuyo papel neurobiológico es integrar la motivación y la acción motora. Su función consiste en transmitir aquella información motivacional relevante que hace que se pongan en marcha las acciones motoras necesarias para lograr la satisfacción o recompensa proyectada.
El centro de recompensa (también conocido como vía mesolímbica), del cual forma parte, nace en el tallo cerebral ―en el área tegmental ventral― en donde se libera el neurotransmisor dopamina que llega hasta el núcleo accumbens.
la motivación del sujeto y traducirla en una acción motora con el fin de cumplir con los objetivos del organismo, es de las principales funciones del núcleo accumbens la de transmitir la información sobre sus conexiones tanto con el prefrontal como con los ganglios basales. Así, permite que hagamos conductas instrumentales, dirigidas a un fin específico.
Esta función y sus conexiones con amígdala cerebral tiene, que ver con un tipo de memoria muy importante: la memoria emocional. Esta capacidad está en la frontera entre los procesos mentales ligados a la emoción y los procesos psicológicos superiores, ya que por un lado trabaja con emociones y por el otro influye en la toma de decisiones y en la creación de conceptos.
2. Influye en la planificación de la conducta
Las conexiones del núcleo accumbens con el con el lóbulo frontal han permitido ver cómo esta estructura tiene participación en la ideación y planificación de la conducta, siendo como hemos dicho un importante punto de integración entre los aspectos motivacionales de la conducta y su puesta en marcha.
3. Evaluación de la situación
La participación de esta estructura también se da nivel evaluativo, al integrar la información emocional con la valoración adaptativa que realiza el frontal. De este modo se consigue asociar un estímulo a una valoración subjetiva mediante un proceso que también tiene que ver con la memoria emocional.
4. Papel en la adicción
El núcleo accumbens juega un importante papel en el proceso adictivo, ya que se encuentra vinculado a la experimentación de recompensa. Este núcleo cerebral forma parte de la vía mesolímbica, formando parte del centro de recompensa cerebral. Concretamente es en esta zona donde las drogas estimulantes actúan, produciendo un aumento en los niveles de dopamina cerebrales.
5. Obtención de placer
Si bien no es la única estructura cerebral vinculada a la experimentación del placer, el núcleo accumbens si mantiene una estrecha vinculación con su consecución. Y es que diferentes experimentos han demostrado que si bien su inhibición no elimina el deseo de obtener un reforzador, sí produce una disminución o supresión de las conductas necesarias para obtener el objeto de deseo. Los datos observados demuestran que la participación del núcleo accumbens se da en procesos adictivos, así como también en la alimentación y el sexo.
6. Aprendizaje y memoria
Los anteriores puntos citados hacen ver que el núcleo accumbens tiene una gran relevancia a la hora de establecer la automatización y aprendizaje de conductas dirigidas a la obtención de una recompensa. También participa en el proceso de habituación.
7. Agresividad y conductas arriesgadas
Una hiperactividad en el núcleo accumbens puede llegar a causar conductas agresivas. Ante una presencia muy elevada de dopamina y otras alteraciones que dificulten la inhibición conductual, puede llegar a producir que se busque la satisfacción personal sin valorar los riesgos.
De hecho, los estudios realizados en personas que tienen psicopatía parecen indicar que estas personas tienen, entre otras alteraciones, un severo desequilibrio en el núcleo accumbens, padeciendo una hiperreactividad a la dopamina que podría inducir a buscar la propia recompensa con indiferencia a las consecuencias para los otros.
Intervienen en procesos emocionales y en clasificar las sensaciones que percibimos solo es valido si se asocia a las múltiples conexiones que tienen con iencefalo y telencefalo.
.
.
Referencias bibliográficas:
Fernández-Espejo, E. (2000). ¿Cómo funciona el nucleus accumbens?. Rev. Neurol. 30: 845-9.
Kandel, E. R. (2001). Principios de Neurociencia. 1º edición. McGraw-Hill.
Salamone, J.D.; Correa, M.; Mingote, S. & Weber, S.M. (2003). Nucleus Accumbens Dopamine and the Regulation of Effort in Food-Seeking Behavior: Implications for Studies of Natural Motivation, Psychiatry and Drug Abuse. Journal of Pharmacology and Experimental Therapeutics, 305 (1). 1-8.
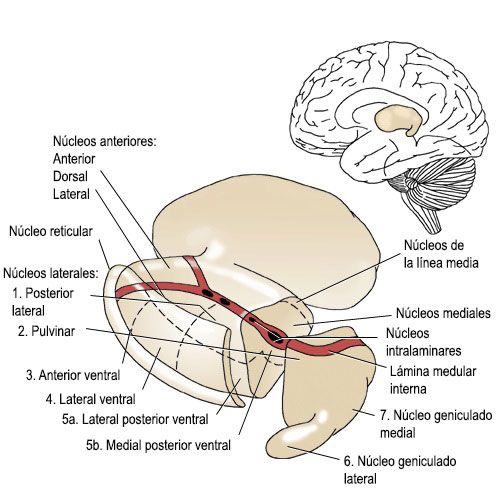
El Tálamo ha sido muy estudiado a lo largo de la historia y recientemente pese a los modernos mecanismos que poseemos sigue siendo una incógnita porque no es producto de una evolución resiente, es antigua forman PARTE del CEREBRO DE LOS REPTILES de forma qué la parte más antigua, se localiza en el núcleos reticulares e intralaminares y cuando se interrumpen desconectan la emoción del sentimiento , como se deduce de los trabajos de Enrique Rubio Una función antigua y otra mas diferenciada que analiza las emociones, el sentimiento
El término tálamo deriva de la palabra griega “thalamos” que significa “cámara interna” o“lecho nupcial”. Galeno (130-200 d. C.)
El sistema nervioso central (SNC) esta integrado por dos grandes regiones, el encéfalo y la médula espinal.
El encéfalo, constituido por todas las estructuras del SNC situadas dentro del cráneo, está formado por el cerebro (hemisferios cerebrales y diencéfalo) y el tronco encefálico (mesencéfalo, protuberancia /cerebelo y bulbo raquídeo).
El diencéfalo está constituido por un conjunto de estructuras nerviosas situadas alrededor del tercer ventrículo y recubiertas en gran parte por los hemisferios cerebrales. Macroscópicamente puede verse en la región basal del encéfalo donde podemos localizar las estructuras diencefálicas inferiores, las que forman el suelo del diencéfalo: quiasma óptico, hipófisis y cuerpo mamilar. En contacto directo con la base del cráneo, sólo separado del hueso por las meninges y espacios entre membranas. En la parte más caudal el diencéfalo se continúa con estructuras mesencefálicas y a derecha e izquierda con los hemisferios cerebrales.
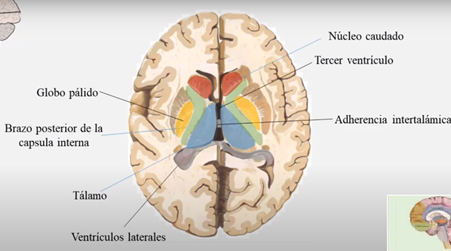
El tálamo
La estructura mayor del diendcefalo compuesta por dos ovoides unidos por una comisura.
Las caras mediales de ambos tálamos están unidas entre sí por un conjunto de fibras intertalámicas conocidas con el nombre de masa intermedia, adherencia intertalámica o comisura gris intertalámica. La cara lateral de forma cóncava, está separada del núcleo lenticular por el brazo posterior de la cápsula interna.
Neuronas del Tálamo.
En el tálamo existen dos tipos de neuronas desde un punto de vista funcional:
1.- neuronas principales o de proyección (transmiten información fuera del tálamo), las cuales representan cerca del 75% de la población neuronal total; y
2.- interneuronas locales o de circuitos locales (pueden recibir información de las mismas fuentes que las neuronas principales pero sólo entran en contacto con células talámicas que participan en la misma etapa de procesamiento), las cuales constituyen alrededor del 25%.
Las neuronas principales envían sus axones a la corteza cerebral, donde liberan un neurotransmisor excitatorio (glutamato generalmente) para activar las neuronas corticales. El glutamato y el aspartato son neurotransmisores excitatorios presentes en las terminaciones corticotalámicas y cerebelosas y en las neuronas de proyección talamocortical. Una excepción lo constituyen las aferencias subcorticales de los núcleos grises de la base que son GABA-érgicas, inhibitorias.
Las neuronas de circuitos locales liberan ácido gammaaminobutírico (GABA) en las células de proyección para inhibirlas. Este neurotransmisor inhibitorio se localiza en las terminaciones que provienen del globo pálido, en las neuronas de circuitos locales y en las de proyección del núcleo reticular y cuerpo geniculado lateral. Son proyecciones GABA-érgicas las principales proyecciones del segmento palidal medial hacia el ventral anterior (parvocelular) y el ventral lateral (pars oralis) y las proyecciones de la parte reticular de la sustancia negra al núcleo ventral anterior (magnocelular) y dorsomedial (paralaminar). Estas aferencias desempeñan un papel fundamental en la función motora [2]. Las neuronas GABA-érgicas han sido identificadas en todas las láminas del cuerpo geniculado lateral, siendo más abundantes en las láminas 1 y 2 (magnocelulares).
Las aferencias procedentes de regiones subcorticales y de la corteza cerebral que se dirigen hasta los núcleos talámicos, excitan (despolarizan) a las neuronas de proyección e interneuronas locales de dichos núcleos. A su vez las neuronas de circuitos locales inhiben (hiperpolarizan) a las neuronas de proyección, el neurotransmisor utilizado es el GABA. Así las aferencias hacia el tálamo influyen sobre las neuronas de proyección (tálamocorticales) a través de dos vías: una excitatoria directa y una inhibitoria indirecta (por medio de las neuronas de circuitos locales). Las neuronas de circuitos locales modulan la actividad de las neuronas de proyección, las cuales envían sus axones a los destinos extratalámicos. Además las células de proyección envían una rama colateral a las neuronas del núcleo reticular talámico, el cual contiene el neurotransmisor inhibitorio GABA, actuando como neuronas de circuitos locales. Las células del núcleo reticular talámico envían ramas axónicas a las neuronas de proyección y de circuitos locales por lo que ambas son inhibidas. La corteza cerebral, la cual recibió proyecciones aferentes excitatorias de las células talámicas de proyección, envía axones excitatorios de regreso a todos los tipos celulares talámicos, por lo que las aferencias corticales activan a las neuronas de proyección así como a las inhibitorias de circuitos locales y del núcleo reticular.
De esta forma el tálamo no sólo es un simple relevo de información entre los centros aferentes y la corteza, sino que es el encargado del procesamiento de la información, influyendo por tanto sobre las funciones corticales [3].
Grupos nucleares talámicos
En el tálamo,se han identificado hasta 50 núcleos talámicos [4], varios de los cuales son subdivisiones microscópicas. La nomenclatura de los núcleos talámicos es muy compleja y en algunos casos se desconocen sus conexiones y la significación funcional de los más pequeños [2].
Su complejidad ha permitido diferentes formas de clasificar sus núcleos y las funciones y pproyecciones de estos.
I- CLASIFICACIÓN DESDE UNA PERSPECTIVA EVOLUTIVA. II- CLASIFICACIÓN ANATOMO-FUNCIONAL. III- CLASIFICACIÓN BASADA EN CRITERIOS CITOARQUITECTÓNICOS. IV- CLASIFICACIÓN TENIENDO EN CUENTA LAS CONEXIONES. V- CLASIFICACIÓN BASADA EN LAS CARACTERÍSTICAS COMPARTIDAS DE CONECTIVIDAD DE FIBRAS Y FUNCIONES.
Desde una perspectiva evolutiva, los Núcleos de la línea media tienen escaso desarrollo en seres humanos y son difíciles de delimitar, localizados en la sustancia gris periventricular, por encima del surco hipotalámico. Mantiene estrechas relaciones con el hipotálamo, los núcleos intralaminares y el núcleo dorsomedial .
Entre los núcleos de la línea a destacar el: – Núcleo paratenial – Núcleo paraventricular – Núcleo reuniens – Núcleo romboide
Participan en las emociones, la memoria y la funciones autonómica.
Núcleos intralaminares Constituyen una numerosa serie de acúmulos neuronales situados en el espesor de la lámina medular interna del tálamo (figura 3). Los dos núcleos principales desde el punto de vista funcional en humanos son el: – Núcleo centromediano – Núcleo parafascicular Otros núcleos que se localizan más hacia la región rostral incluye el: – Núcleo paracentral – Núcleo central lateral
– Núcleo central medial.
Los núcleos intralaminares, elementos que funcionan, llevando y modulando sensibilidades plurales inespesificas muy primitivas
Los núcleos intralaminares influyen sobre la actividad cortical a través de otros núcleos talámicos. Desempeñan una función global activadora, debida a sus conexiones múltiples extratalámicas y corticales y una función específica. Las conexiones que estos núcleos mantienen con el putamen y el caudado contribuyen al control motor subcortical.
Núcleos reticulares Recubren el polo anterior y la cara lateral del tálamo, del que queda separado por una delgada hoja de sustancia blanca, la lámina medular externa. Se localizan entre la lámina medular externa y la cápsula interna
Entre estos núcleos reticulares destacamos los siguientes: – núcleo reticular. es una delgada lámina de sustancia gris vertical, que se apoya sobre la cara externa del tálamo. – núcleos reticulares de la línea media. pequeñas masas grises adosadas a la cara medial del tálamo. – núcleo centro mediano. es un núcleo grande situado en el espesor de la lámina medular externa. los núcleos intralaminares, reticular y de la línea media considerados habitualmente como inespecíficos, están relacionados con el DESPERTAR, CONTROL MOTOR Y LA CONCIENCIA DE LAS EXPERIENCIAS SENSORIALES.
CLASIFICACIÓN TENIENDO EN CUENTA LAS CONEXIONES
Se diferencian dos grandes grupos nucleares talámicos: específicos e inespecíficos.
1.- Núcleos específicos: aquellos que tienen una relación específica y selectiva con una parte concreta de la neocorteza. Es decir, son eslabones intermedios en el procesamiento de la información hasta la corteza y funcionan de un modo complejo, colaborando en la integración, selección, procesamiento y transmisión hacia el cortéx cerebral. Cada uno de estos núcleos recibe proyecciones desde el área de la corteza cerebral a la que ha enviado sus eferencias, es decir existe una reciprocidad entre estos núcleos y la corteza cerebral. Se conocen también como núcleos corticodependientes. Dentro de estos núcleos específicos se diferencian dos tipos: – Núcleos específicos de relevo: reciben aferencias directas de las áreas subcorticales y se proyectan a la capa IV de la corteza cerebral. Este grupo lo integran los siguientes núcleos: grupo nuclear anterior, núcleo VA, núcleo VL, VP, CGL y CGM. Según Martín [10] los núcleos de relevo son esenciales para todas las funciones cerebrales, y cada uno de ellos desempeña un papel diferente en la percepción, la volición o la cognición, transmitiendo información desde estructuras subcorticales concretas a una porción limitada del córtex. – Núcleos específicos de asociación: reciben aferencias corticales y de otros núcleos talámicos y a su vez ellos se proyectan a diversas capas corticales (I, III y VI). Este grupo lo integran los siguientes núcleos: DL, LP, DM y pulvinar.
2.- Núcleos inespecíficos: se proyectan a varias regiones corticales y subcorticales. Son los núcleos de la línea media, intralaminares y reticulares. Para estos núcleos la corteza cerebral no es su lugar de proyección principal. Se ha comprobado que los dos primeros envían proyecciones a regiones subcorticales precisas, por lo que no son tan inespecíficos como se pensaba [2]. Actúan en el despertar y regulando la excitabilidad de las regiones más amplias de la corteza cerebral [10]. Se le denomina también núcleos corticoindependientes. V- Clasificación basada en las características compartidas de conectividad de fibras y funciones Según Afifi y Bergman [6] en general se utilizan dos sistema de nomenclatura:
A).- Núcleos talámicos agrupados en tres categorías generales teniendo en cuenta la conectividad de las fibras. 1.- De modalidad específica: comparten las siguientes características: a.- reciben aferencias directas de tractos ascendentes largos relacionados con información somatosensorial, visual y auditiva (VPL y VPM, CGL y CGM) o cualquier proceso de información derivado de los núcleos grises de la base (VA, VL), el cerebelo (VL) o el sistema límbico (núcleo anterior y DL). b.- tienen conexiones recíprocas con áreas corticales bien definidas (área somatosensorial primaria, auditiva y visual, áreas premotoras y motoras primaria y giro cingulado) c.- Se degeneran mediante la ablación de las áreas corticales específicas a las que se proyectan. 2.- Multimodal asociativo: no reciben aferencias directas de los tractos ascedentes largos y se proyectan áreas de asociación en los lóbulos frontales, parietal y temporal. Incluyen, el DM y el complejo nuclear pulvinar-lateral posterior. Tienen la siguientes características en común: 1- No reciben aferencias de los tractos ascendentes largos; 2- la mayor parte de sus aferencias provienen de otros núcleos talámicos; 3- su proyección principal se dirige a las áreas de asociación de la corteza cerebral. 3.- Inespecíficos y reticular: se caracterizan por proyecciones corticales indirectas difusas y amplias y por aferencias de la formación reticular troncoencefálica. Incluyen los núcleos intralaminares, de la línea media y reticulares.
B).- Teniendo en cuenta la función que desempeñan 1.- Motores: reciben aferencia motoras de los núcleos grises de la base (VA) o el cerebelo (VL) y se proyecta a las cortezas premotoras y motoras primarias. 2.- Sensitivos: recibe aferencias de los sistemas ascendentes somatosensoriales (VPL y VPM), auditivo (CGM) y visual (CGL). 3.- Límbicos: se relacionan con estructuras límbicas (cuerpos mamilares, hipocampo, giro del cíngulo). 4.- Asociativos: se corresponden con los núcleos multimodales asociativos. 5.- Inespecífico y reticular: corresponden a la misma categoría del otro sistema de nomenclatura. Conexiones tálamo-corticales y cortico-talámicas
La organización en el seno de la corteza cerebral de las proyecciones tálamo-corticales y cortico-talámicas y las propiedades neurofisiológicas de las fibras que ascienden o descienden a o desde la corteza cerebral son la base de las complejas relaciones entre los diferentes núcleos talámicos y la corteza cerebral [11].
Fue Lorente de Nó [12] quien describió las aferencias tálamo-corticales como: fibras tálamo-corticales específicas y fibras tálamo-corticales inespecíficas. Las primeras tienen su origen en los núcleos específicos del tálamo, sinaptan en la capa IV de la corteza y son portadoras de información de la sensibilidad general y especial (excepto la olfativa). Las segundas son fibras ascendentes con colaterales fundamentalmente a las capas I, II y VI. Estas vías inespecíficas están relacionadas con las vías tálamo-corticales difusas, procedentes de los núcleos de la línea media e intralaminares hacia el córtex cerebral [13,14] y relacionados con los mecanismos de arousal.
Macchi [13] en estudios realizados en gatos señala que existen cuatro tipos de conexiones tálamo-corticales, en función de la difusión de sus eferencias.
La primera incluye todos los núcleos que envían sus proyecciones sobre un área anatomofuncional homogénea de la corteza cerebral, según una topografía ordenada, como es el caso de los núcleos VPL y VPM y el núcleo DM. La segunda categoría viene dada por todos los núcleos cuyas eferencias fundamentalmente se dirigen a áreas corticales funcionalmente homogéneas, proyectándose también sobre otras áreas corticales alejadas de las primeras, aunque funcionalmente iguales. Se incluirían en esta categoría el LP y el pulvinar.
La tercera categoría engloba aquellos núcleos que envían sus eferencias a varias áreas corticales funcionalmente diferentes. Pertenecen a esta categoría los núcleos intralaminares, VL y VA. Por ejemplo hacia la corteza motora y hacia la corteza límbica.
La última categoría, incluye a los núcleos talámicos que se proyectan de manera difusa, en zonas corticales sensoriales y perisensoriales somatoauditivas. El complejo posterior y en particular por el CGMmc son los representantes de esta categoría.
Hemos de señalar también que existen proyecciones recíprocas de todos los núcleos de relevo y de algunos núcleos de asociación que van desde el tálamo a la corteza y desde la corteza al tálamo a través de la cápsula interna, denominadas radiaciones talámicas. Tales fibras irradian desde y hacia el tálamo como si fuera un abanico dando así lugar a la corona radiada. A pesar de que estas radiaciones establecen conexiones prácticamente con todas las partes de la corteza, la riqueza de las conexiones varia en las diferentes áreas corticales. Las más abundantes son hacia la circunvolución precentral y postcentral, área calcarina circunvolución de Heschl, región parietal posterior y partes adyacentes del lóbulo temporal [2]. Se agrupan en cuatro grupos de fibras o pedúnculos talámicos:
1.- Pedúnculo anterior, asciende hacia el lóbulo frontal por el brazo anterior de la cápsula interna. Constituido por la proyecciones del núcleo anterior del tálamo con el cíngulo y del DM con la corteza prefrontal. 2.- Pedúnculo superior, finaliza en la corteza parietal tras recorrer la porción lenticulotalámica de la cápsula interna. Formado por la fibras del núcleo VP y los núcleos VA y VL. 3.- Pedúnculo inferior, ocupa la porción sublenticular de la cápsula interna y contiene la radiación auditiva que finaliza en la corteza auditiva. 4.- Pedúnculo posterior, a través de la porción retrolenticular del brazo posterior de la cápsula interna se dirige al lóbulo occipital. Lo integran las proyecciones o radiaciones ópticas del CGL y del complejo nuclear talámico lateral posterior-pulvinar. Conexiones de los núcleos talámicos
A continuación analizaremos las principales aferencias que llegan a los diferentes núcleos talámicos y las eferencias que de ellos parten. Utilizaremos para este fin la clasificación anatomo-funcional mencionada anteriormente.
1.- Grupo nuclear anterior Las principales aferencias a este grupo nuclear proceden del hipotálamo (cuerpos mamilares) y formación hipocámpica. El tracto mamilo-talámico está topográficamente organizado, de modo que el núcleo mamilar medial se proyecta ventralmente al AV, AM y el lateral lo hace al AD. Las fibras de la mitad lateral del tracto, que comprende todas las fibras procedentes del núcleo mamilar lateral y la mitad lateral del medial, se proyectan de forma bilateral. El resto son homolaterales. Las principales eferencias las envía hacia la circunvolución cingulada (corteza asociativa límbica). También envía sus axones a la corteza prefrontal, corteza motora medial, áreas orbitarias de la corteza frontal y áreas de asociación visual.
2.- Núcleo dorsomedial Las aferencias del núcleo DM proceden del bulbo olfatorio (DMmc), la amigdala (DMmc), sustancia negra (DMpl), pálido (DMmc, DMpc), TCS (DMpl), núcleos talámicos intralaminares y de la hilera dorsal. Las conexiones del DM-córtex prefrontal y el córtex frontal-DM son extitatorias -glutaérgicas-, mientras que las aferencias procedentes del pálido y sustancia negra son inhibitorias -GABA-érgicas- [15]. El DM estable conexiones recíprocas con diversas áreas corticales donde sus fibras acaban en la capa I, III y VI. Entre ellas destacan la corteza prefrontal e insular (DMmc, DMpc), la corteza de asociación (DMmc) y la corteza del campo ocular (área 8) (DMpl).
GRUPO NUCLEAR LATERAL
ZONA DORSAL El núcleo DL recibe impulsos del hipocampo (a través del fórnix) tal vez de los cuerpos mamilares y áreas asociativas de la corteza parietal. Se proyecta al giro del cíngulo y zonas asociativas del lóbulo parietal. No se conocen bien las aferencias del LP, pero parece ser que proceden de los núcleos de relevo adyacentes en especial del núcleo VP, CGL del TCS y de la corteza sensorial secundaria (área 2) con la que establece conexiones recíprocas. Los estudios de transporte retrógrado indican que el LP emite sus eferencias a las áreas asociativas 5 y 7 de la corteza parietal [2]. Se proyecta también a la corteza visual (áreas 17, 18 y 19).
Zona ventral El núcleo VA recibe aferencias fundamentalmente del núcleo dentado del cerebelo, del globo pálido y de las porciones reticulares de la sustancia negra y áreas no motoras como el TCS. Hasta él llegan también fibras procedentes de los núcleos intralaminares y de la línea media. Las eferencias se dirigen fundamentalmente hacia los núcleos intralaminares, áreas 6 y 8 y áreas orbitofrontales.
Las aferencias del núcleo VL proceden fundamentalmente de los núcleos cerebelos profundos. El sistema dentotalámico constituye la principal aferencia. Aunque el sistema de fibras pálido-talámicas se proyectan en su mayor parte sobre las neuronas del núcleo VA algunas fibras alcanzan el núcleo VL. Hasta esta zona talámica llegan también aferencias procedentes de la corteza prefrontal, motora primaria (área 4) y núcleos intralaminares. Este núcleo envía sus eferencias hacia la corteza motora primaria, premotora y motora suplementaria y áreas somatosensitivas no primarias en la corteza parietal (áreas 5 y 7).
Los núcleos cerebelosos profundos se proyectan fundamentalmente al VL y desde el pálido preferentemente al VA. El núcleo VP constituye el principal destino de las fibras del lemnisco medio, lemnisco trigeminal y el tracto espino-talámico. La parte oral del núcleo VPLo recibe proyecciones de los núcleos cerebelosos profundos contralaterales y se proyecta a la corteza motora primaria. La parte caudalis, VPLc recibe aferencias somatosensoriales de la médula espinal y núcleos del bulbo raquídeo a través del lemnisco medial y haces espinotalámicos.
El VPM recibe aferencias somáticas de receptores de la cara y estructuras intraorales. Las fibras trigeminales ascendentes llegan hasta este núcleo.
Ambos núcleos, VPL y VPM también reciben aferencias de la corteza somatosensorial primaria. Sus eferencias poseen una precisa proyección tópica a la corteza de la circunvolución postcentral, áreas 3, 1 y 2 (hómunculo sensitivo).
4.- Grupo nuclear posterior El pulvinar no recibe proyecciones de las vías sensitivas largas ascendentes, excepto la parte inferior de este núcleo a la cual llega una proyección de las capas superficiales de los TCS. La parte inferior de este núcleo y la porción adyacente del pulvinar lateral mantiene conexiones recíprocas con la corteza occipital incluida la corteza estriada. Las proyecciones del pulvinar inferior a las áreas 17,18 y 19 constituyen el enlace final en una vía visual extrageniculada.
La parte lateral del pulvinar se proyecta a la corteza temporal y recibe proyecciones recíprocas de la misma región. La parte medial se proyecta a la circunvolución temporal superior.
El complejo pulvinar-lateral posterior y el núcleo DM se conocen en conjunto como núcleos talámicos multimodales de asociación. Se pueden considerar conjuntamente tanto desde el punto de vista de sus conexiones como de sus funciones. Reciben proyecciones desde la retina y TCS, se relacionan con la corteza P-T-O asociativa y también con áreas visuales de la corteza cerebral. Desde un punto de vista funcional se le relaciona con la vía visual y también con el control de movimientos oculares.
La parte ventral del CGM recibe las proyecciones del núcleo central del tubérculo cuadrigémino inferior (TCI) y sus eferencias se dirigen hacia la corteza auditiva primaria (área 41). Esta proyección está tonotópicamente organizada de manera que las frecuencias bajas se sitúan lateralmente y las frecuencias altas lo hacen medialmente. A la región dorsal, llegan aferencias fundamentalmente procedentes del núcleo pericentral del TCI, su organización tonotópica no es tan clara como la de la región ventral y se proyecta a la corteza auditiva secundaria (áreas 22 y 42). La región medial recibe las proyecciones del núcleo externo del TCI y se proyecta sin tonotopía a toda la corteza auditiva (áreas 22, 41 y 42).
El CGL recibe proyecciones de la retina por la cintílla óptica y se proyecta a la corteza calcarina (área 17) a través del haz geniculocalcarino o radiación visual y recibe fibras corticogeniculadas del mismo área. En menor medida se proyecta a las áreas asociativas visuales adyacentes. Establece conexiones internucleares con el pulvinar. Las proyecciones retinogeniculadas estan tópicamente organizadas. Los cuadrantes superiores de ambas retinas (campo visual inferior) finalizan en la mitad superomedial del CGL. Los cuadrantes inferiores (campo visual superior) lo hacen en la mitad inferolateral del núcleo.
NÚCLEOS DE LA LÍNEA MEDIA Las principales aferencias hacia estos núcleos proceden del hipotálamo, núcleos del tronco encefálico, amigdala y giro parahipocampal. Envían sus proyecciones hacia la corteza límbica y el núcleo estriado ventral. NÚCLEOS INTRALAMINARES Se consideran clásicamente y en conjunto como núcleos que reciben y envían proyecciones de modo inespecífico o difuso. No obstante también es cierto que diferentes porciones de los mismos reciben aferencias distintas y cada una de ellas proyecta de modo topográfico preciso a los ganglios basales y a zonas concretas de la corteza cerebral, no siendo tan inespecíficos como se creía [2]. Las principales aferencias hacia estos núcleos proceden de la formación reticular. El sistema dentorrubrotalámico (cerebelo) se proyecta fundamentalmente al VL y colaterales de este sistema se proyectan a los núcleos intralaminares. La proyección principal de las fibras palidotalámicas es el núcleo VA, colatarerales de esta proyección alcanzan los núcleos intralaminares. La mayor parte de las fibras del tracto espinotalámico y lemnisco trigeminal (fibras ascendentes de las vías del dolor) se proyectan sobre el VP, pero también a los núcleos intralaminares.
Estos núcleos reciben proyecciones también de fibras corticales procedentes de las áreas motoras y premotora. Las fibras que se originan en la corteza motora (área 4) terminan en las neuronas de los núcleos centromediano, paracentral y centrolateral. Las originadas en la corteza premotora (área 6) concluyen en los núcleos parafascicular y centrolateral. En contraste con otros núcleos talámicos, las conexiones entre los núcleos intralaminares y la corteza cerebral no son recíprocas. Las principales eferencias de los núcleos intralaminares se dirigen a otros núcleos talámicos (fundamentalmente reticulares), estriado (caudado y putamen) y ampliamente sobre áreas asociativas de la corteza cerebral. Por métodos de marcaje retrógrado se ha visto que un pequeño número de neuronas intralaminares, proyectan por colaterales axónicos en áreas corticales bastantes alejadas entre si [16]. 7.- Núcleos reticulares Recibe proyecciones cortico-talámicas, pero carece de proyecciones tálamo-corticales. Se encuentran conectados con muchos núcleos talámicos y además son atravesados, recibiendo colaterales, de las fibras tálamo-corticales y cortico-talámicas, ejerciendo por tanto una acción moduladora sobre la actividad neuronal talámica.
ASPECTOS FUNCIONALES
Incidencia del tálamo en los procesos psicofuncionales básicos: sensitivo/motor
El tálamo junto con la corteza cerebral juegan un papel importante en el análisis e integración de las funciones sensitivas. Toda la información sensorial excepto la olfativa (la información se transmite directamente a la corteza temporal medial) se dirigen al tálamo donde hacen escala y se proyectan a las correspondientes áreas corticales específicas.
El CGM está relacionado con la vía auditiva. El input es bilateral, aunque predominan las aferencias del oido opuesto. Las aferencias de este núcleo, se dirigen hacia las áreas auditivas 41 y 42 (áreas auditivas primaria y secundaria auditiva, respectivamente) y hacia el complejo talámico asociativo dorso-pulvinar, de cual salen eferencias hacia las áreas 21 (área inferotemporal visual, circunvolución temporal, relacionada con la visión de la forma) y 22 (corteza auditiva superior. Área de Wernicke) de la corteza cerebral.
El tálamo está implicado también en los mecanismos de la visión. Las aferencias procedentes de la retina terminan en el CGL. Las eferencias se dirigen hacia la corteza visual (área 17) y hacia el complejo asociativo dorso-pulvinar para proyectarse hacia las áreas 18 (corteza visual primaria), 19 (visual secundaría), 1b (somatosensorial primaria), 39 (asociativa parieto-temporo occipital) y 37 (asociativa parieto-temporo-occipital) de la corteza cerebral.
El tálamo forma parte del sistema somatosensitivo colaborando en la percepción de estímulos mecánicos, térmicos y dolorosos. El núcleo VP recibe los tractos ascendentes largos que conducen las modalidades sensoriales, incluso del gusto, de la mitad contralateral del cuerpo y la cara. Este núcleo envía eferencias al pulvinar y al núcleo LP. Estos a su vez envían eferencias a la corteza parietal zonas esta relacionadas con el reconocimiento somatoestésico.
Gracias a las proyecciones del VP hacia las áreas 5 (corteza sensorial somestésica terciaría, área asociativa parietal posterior), 7 (áreas asociativa parietal posterior, relacionada con la percepción visuo-motora) y área 40 (asociativa parieto-temporo-occipital) es posible llevar a cabo funciones como es el reconocimiento de los objetos por el tacto (esterognosia) y del propio cuerpo (somatognosia).
El VPL actúa como relevo para la información somática del cuerpo y las extremidades, ya que dirige sus proyecciones hacia la corteza somestésica primaria en la circunvolución postcentral (área 3, 1, 2,) en la cual se analiza la información sensitiva cutánea, muscular, tendinosa, articular y visceral, siendo posible de esta manera las percepciones objetivas como la forma, el tamaño, la textura, la temperatura y el peso.
El VPM sirve de centro de relevo sensitivo-talámico de la cabeza y cara. Las eferencias de este núcleo se dirigen a través de la cápsula interna hasta la corteza somestésica primaria del lóbulo parietal. A través de la proyecciones de esta zona talámica hacia zonas frontales (áreas 4, 8, 6, 44 y 45) el tálamo está involucrado en la sensopercepción de los movimientos.
El tálamo está implicado también en los mecanismos del dolor. Los principales núcleos de destino de los axones ascendentes para el dolor y la temperatura, se encuentran el núcleo VP. El VPM y VPL reciben la mayor parte de estas aferencias. El VPM reciben información nociceptiva desde la cara y el VPL del resto del cuerpo. La disposición similar de los estímulos mecanosensitivos y nocivos es la responsable de los mecanismos discriminativos del dolor [19].
Los núcleos talámicos intralaminares en cuanto al dolor se refiere participan en la evocación de la respuesta desencadenada por un estímulo nocivo a través de las proyecciones que llegan a estos núcleos desde la formación reticular.
Algunas modalidades sensitivas se perciben a nivel talámico, hecho este que se pone de manifiesto cuando existen lesiones o ablaciones de la corteza cerebral. En estos casos tras la lesión se pierde toda la sensibilidad contralateral a la lesión recuperándose el dolor, la temperatura y la sensibilidad epicrítica (burda). En la clínica está bien descrito este cuadro, conocido como síndrome talámico. En estos casos el umbral de estimulación que producen estas sensaciones es elevado, las modalidades sensoriales son exageradas y displacenteras, además se suelen acompañar con una marcada respuesta afectiva, normalmente atribuible a la indemnidad del núcleo DM (frecuente en lesiones vasculares).
Lesiones vasculares que afectan al territorio talámico postero-lateral (núcleos VPL, VPM, CGM, pulvinar y centromediano) pueden dar lugar a pérdida sensorial contralateral, paréstesias y dolor talámico. Bien descrito es el síndrome de Dejerine y Roussy, caracterizado por un dolor intenso, persistente y paroxístico a menudo intolerable, que se suele presentar en el momento de la lesión o después de un periodo de hemiparesia transitoria, hemiataxia y pérdida sensitiva hemicorporal.
Por otro lado la participación del tálamo en el control motor queda reflejado por las aferencias procedentes de núcleos grises de la base, cerebelo y corteza motora que llegan a él y las eferencias que de él parte hacia la corteza motora y premotora. En el sistema motor intervendran fundamentalmente los siguientes núcleos: VA y VL, núcleos intralaminares y núcleos reticulares y podemos destacar dos grandes sistemas: palidal y cerebeloso. La separación entre ambos circuitos es debido a que las aferencias son distintas y también sus eferencias hacia áreas corticales en donde proyectan. Alteraciones en las proyecciones del VL pueden dar lugar a trastornos motores (discinesias). Lesiones en este núcleo disminuyen los movimientos anormales cerebelosos y de los núcleos grises de la base [6].
Lesiones en el núcleo ventral intermedio (Vim), núcleos ventrales caudales, centromediano, núcleos sensoriales y pulvinar pueden causar gran variedad de alteraciones del movimiento, incluyendo, distonias, temblor, balismo, corea, entre otros [20-22]. Lesiones vasculares que afectan a los núcleos VA, VL, DM y núcleo anterior pueden causar hemiparesia contralateral y trastornos de los campos visuales.
Existe evidencia de que los núcleos intralaminares también están implicados en el control de los movimientos. Este núcleo recibe aferencias principalmente de la formación reticular, del pálido, putamen, núcleos subtálamicos y de áreas corticales (6 y 4). Las conexiones que estos núcleos mantienen con el putamen y el caudado, contribuyen al control motor subcortical. El núcleo centromediano recibe aferencias del pálido, sustancia negra (zona reticular), zona incierta, núcleos profundos del cerebelo, córtex motor primario y núcleos reticulares [23,24]. Envía amplias proyecciones glutamato-érgicas excitatorias al putamen y proyecciones difusas al borde dorsolateral del núcleo caudado y núcleos subtalámicos [25, 26]. Los núcleos reticulares talámicos, terminan de manera difusa en la corteza cerebral, permitiendo la activación necesaria para el correcto funcionamiento del sistema motor. Existen trabajos que señalan cierta implicación de los núcleos de la línea media con el sistema motor. Lee y Marsden [21] que señalan las lesiones causantes de las distonías tálamicas no hay que situarlas en los núcleos VA y VL sino en zonas más posteriores o en los núcleos de la línea media. Podemos describir una semiología motriz que caracterizaría a las lesiones talámicas: 1.- alteraciones del sistema motor voluntario (incoordinación cerebelosa contralateral, sincinesias homolaterales de imitación y contracturas); 2.- alteraciones del sistema motor involuntario; 3- perturbaciones globales del movimiento (mano talámica, caracterizada por movimientos incesantes de los dedos, tanto en el plano horizontal como en el vertical); 4.- alteraciones de la marcha [27]. Incidencia del tálamo en los procesos psicofuncionales superiores: nivel atencional/ emoción/ lenguaje/ memoria/ función ejecutiva El tálamo regula funciones de la corteza asociativa y es importante en funciones como el lenguaje, el habla y funciones cognitivas, mediadas corticalmente [28]. Hay tres regiones importantes de corteza asociativa: parieto-temporo-occipital, prefontral y límbica, hacia las cuales se proyectan diferentes núcleos talámicos. Así la corteza parieto-temporo-occipital (áreas 39 y 40) está relacionada con funciones perceptivas, visión, lectura y recibe información del pulvinar. La corteza asociativa prefrontal es importante para la planificación de la conducta y los movimientos, cognición, aprendizaje, memoria y pensamiento. El DM proyecta sus fibras hacia esta zona cortical. Un estudio reciente realizado en monos a los cuales se les realizó una ablación del núcleo DMmc, ha puesto de manifiesto que lesiones en esta zona talámica causan trastornos de memoria debidos principalmente a la interrupción de la función entre este núcleo y el cortex prefrontal [29]. La corteza límbica, relacionada con el aprendizaje, la memoria y la emoción, recibe fundamentalmente eferencias del núcleo anterior talámico.
Tálamo y nivel atencional
La participación del tálamo y de la formación reticular en la regulación del nivel de arousal (vigilancia) se puso de manifiesto ya en la primera mitad del siglo XX con los trabajos pioneros realizados por Morison y Dempsey [30], Jasper [31] y Moruzzi y Magoun [32]. Los núcleos intralaminares están relacionados con la excitabilidad general de la corteza cerebral al transmitir información procedente de la formación reticular mesencefálica a múltiples áreas corticales y al cuerpo estriado, desempeñando un papel importante en el control del sueño y la vigilia. La estimulación eléctrica de estos núcleos provoca una activación generalizada de la corteza cerebral (recruiting response) formando parte del sustrato anatómico del sistema reticular activador ascendente y por lo tanto de los mecanismos del sueño y la vigilia.
Los núcleos de la línea media parecen ser el lugar por el que el tálamo, junto con la formación reticular, controla las señales que acceden a la corteza cerebral. Trabajos realizados en este campo, señalan que el tálamo regula el nivel de arousal cortical a través de las conexiones tálamo-corticales que se originan en los núcleos DM, intralaminares y de la línea media y a través de las interacciones intratalámicas con los núcleos reticulares [23,33].
Los estudios llevados a cabo en especies animales han proporcionado evidencias de que los núcleos reticulares están relacionados con el ciclo sueño-vigilia [23,33]. Se ha comprobado que las neuronas GABA-érgicas de los núcleos reticulares controlan la actividad de las neuronas tálamo-corticales, modulando así la actividad cortical [33,34].
En humanos se ha visto en estudios realizados con técnicas de neuroimagen funcional que existen variaciones en el flujo sanguíneo tálamico en función del nivel de conciencia [35,36]. Kinomura, Larsson, Gulyás y Roland [37] han demostrado cambios en el flujo sanguíneo de los núcleos intralaminares del tálamo y la formación reticular en función del nivel de arousal de sujeto.
En una investigación llevada a cabo por Fiset y cols. [38] donde se manipulaba el nivel de conciencia de los sujetos utilizando Propofol (fármaco con propiedades anestésicas que disminuye el flujo sanguíneo cerebral, lo cual se acompaña de una reducción del requerimiento metabólico cerebral de oxígeno y de la disminución de la presión intracraneal), encontraron relación negativa entre el flujo sanguíneo talámico (con PET) y la concentración de propofol utilizada. Los efectos de esta droga anestésica son más pronunciados en la zona medial talámica, el giro cingulado, giro orbitofrontal y giro angular. Parece ser que las variaciones observadas en el tálamo (especialmente en la zona medial) están significativamente relacionadas con la actividad de la formación reticular. Estos autores sugieren que el sistema reticulo-talámico juega un papel fundamental en la modulación de la conciencia.
En la clínica se ha observado que lesiones vasculares en los núcleos intralaminares y dorsomediales pueden causar mutismo acinético y el síndrome de Kleine-Levin (síndrome de hipersomnia y bulimia). Este síndrome se caracteriza por periodos recurrentes de excesiva somnolencia, hiperfagia, hipersexualidad y alteraciones de la memoria reciente.
Diferentes aspectos de la atención pueden ser atribuibles al córtex prelímbico y núcleo DM [39]. Infartos tálamicos pueden causar negligencia y déficits atencionales del espacio extrapersonal contralateral a la lesión [40-42].
Tálamo y emoción
El tálamo interviene en los procesos emocionales y motivacionales. Los principales núcleos implicados son el VA, DM y grupo nuclear anterior.
El VA recibe aferencias desde el cuerpo mamilar y proyecta fibras hacia el cíngulo.
El núcleo DM recibe desde el hipotálamo y la amigdala, enviando sus fibras hacia el lóbulo prefrontal. El DM con sus proyecciones hasta la corteza prefrontal y estructuras límbicas, participa en la integración de la información visceral con el afecto, las emociones y el pensamiento.
El grupo anterior media información visual y emocional. La estimulación eléctrica y la ablación de este núcleo inducen cambios en la tesión arterial y los impulsos motivacionales.
Tálamo y lenguaje
Penfield y Roberts en [43] fueron los primeros en destacar que el tálamo con sus extensas proyecciones corticales está relacionado con funciones linguísticas.
En el lenguaje intervienen fundamentalmente el pulvinar, el grupo nuclear lateral (fundamentalmente el VPL y VPM) y el grupo nuclear anterior. Existen conexiones reciprocas entre el pulvinar y la corteza cerebral importantes para el lenguaje y el pensamiento simbólico (hacia la encrucijada funcional parieto-temporo-occipital). El VPL y VPM participan en el lenguaje gracias a las relaciones que mantienen con áreas somestésicas y a la integración específica que en ellos se produce.
Existe evidencia electrofisiológica de la participación del tálamo en los aspectos motores del lenguaje. Mateer [44] encontró un incremento en la duración de la respuesta verbal después de estimular el tálamo izquierdo dando como resultado una mala pronunciación de las palabras y cambios articulatorios. Posteriormente Bhatnagar y Andy [45] observaron tras la estimulación del núcleo centromediano izquierdo espasmos motores articulatorios.
Johnson y Ojemann [46] señalan que la zona ventro-lateral del tálamo izquierdo (especialmente la parte central) participa en la integración de los mecanismos motores del habla, incluyendo la respiración, ya que tras la estimulación de esta zona talámica se observa una inhibición de la respiración, enlentecimiento del habla y presencia de perseveraciones.
El pulvinar, no está sólo intercalado entre las vías óptica y acústica, sino que proyecta a zonas corticales importantes para el lenguaje y el pensamiento simbólico (encrucijada parieto-temporo-occipital). Lesiones en el núcleo anterior o en el pulvinar pueden causar anomia, parafasias semánticas y errores sintácticos [47].
Ojemann [48] encontró que tras la estimulación de la zona anterior (parte más lateral) del tálamo aparecen repeticiones de palabras que previamente han sido denominadas correctamente. Si la estimulación se realiza en la parte central de la zona ventrolateral aparecían perseveraciones. La estimulación de la parte posterior de la zona ventrolateral y pulvinar anterior daba lugar a la aparición de omisiones y errores en la denominación de objetos.
Tálamo y memoria
En cuanto a la memoria se refiere parece ser que son los núcleos talámicos anteriores, núcleos de la línea media, núcleos dorsomediales y núcleos intralaminares los implicados en los procesos mnésicos, aunque no existen evidencias concluyentes que indiquen cual de estas estructuras es crucial para el buen funcionamiento de la memoria anterógrada [49]. Weiskrantz [50] señala que los déficits de memoria que suelen aparecer en pacientes con lesiones talámicas son similares a los observados tras lesiones en el lóbulo temporal medial: déficits en la codificación de nueva información dando como resultado una alteración en la memoria anterógrada, estando intacta la memoria a corto plazo.
Existe evidencia de alteraciones mnésicas tras lesiones talámicas específicas, en especial en el núcleo DM [51], núcleo anterior [52, 53] y núcleos intralaminares [54].
Parece ser que el núcleo anterior está relacionado con el proceso de consolidación de la información permitiendo la formación de trazos mnésicos y con la memoria de trabajo [55].
Recientemente Celerier, Ognard, Decorte y Beracochea [56] han demostrado en ratones que lesiones en el núcleo anterior causan alteraciones en la ejecución de tareas mnésicas. Según estos autores este grupo nuclear está relacionado con el mantenimiento de la información en el tiempo independientemente de la naturaleza de la información y con los procesos asociativos de la información unimodal y polimodal.
Los núcleos anteriores del tálamo están implicados en los procesos de organización temporal de la memoria [57]. Los núcleos intralaminares permiten la salida de trazos mnésicos ya memorizados, es decir el proceso de activación.
En los procesos de organización temporal de los recuerdos recientes y antiguos intervienen los núcleos DM. Lesiones en estos núcleos pueden dar lugar a una desorganización temporal del recuerdo que afectaría no solo a la información nueva, sino también a la antigua. Pueden aparecer fabulaciones como las descritas en el síndrome de Korsakoff. Victor, Adams y Collins [58] que en los pacientes con síndrome de Korsakoff el núcleo DM está afectado en el 100% de los sujetos, junto con los cuerpos mamilares. El déficit es más severo si están implicados los núcleos DM del tálamo y los núcleos de la línea media [59]. Además, en el síndrome de Korsakoff [60] se ha encontrado relación entre la amnesia anterógrada y el grado de atrofia en los núcleos de la línea media, no evidenciándose relación con la atrofia en los cuerpos mamilares, hipocampo o giro parahipocampal.
Gaffan y Parker [29] en un estudio realizado con monos han encontrado que la parte magnocelular del núcleo DM juega un papel importante en la memoria. Una lesión en esta zona da lugar a una alteración en esta función cognitiva atribuible a la desconexión con el córtex prefrontal.
Sin embargo y a pesar de estos resultados, todavía existe controversia de si lesiones en el DM pueden causar déficits de memoria. En una extensa revisión realizada por Van der Werf y cols [61] sobre los déficits neuropsicológicos que pueden aparecer tras infartos talámicos señalan, que no existe evidencia suficiente para poder establecer la relación del DM con los problemas mnésicos que ocurren después de lesiones diencefálicas. Concluyen que los déficits mnésicos que pueden aparecer y que son compatibles con un “síndrome amnésico”, dependen de la integridad del tracto mamilo-talámico.
La participación del tálamo en el procesamiento mnésico se ha puesto también de manifiesto a través de los estudios electrofisiológicos realizados. Ojemann [48] encontró que la estimulación ventrolateral talámica afecta a la memoria verbal a corto plazo. La estimulación de esta zona durante la presentación del material que posteriormente será evocado reduce el número de errores. La estimulación del pulvinar izquierdo altera el procesamiento mnésico verbal, mientras que la estimulación del pulvinar derecho altera el procesamiento mnésico no verbal [46]. Tálamo y función ejecutiva
Lesiones en el tálamo también pueden causar alteraciones en las funciones ejecutivas, atención, iniciativa, inhibición y organización temporal de la conducta, funciones estas relacionadas con el córtex prefrontal. Se ha propuesto que entre los núcleos talámicos implicados en la función ejecutiva se encuentran el DM, los intralaminares y los núcleos de la línea media. Algunos pacientes muestran deterioro en el funcionamiento ejecutivo después de infartos selectivos del DM [52, 62]. Mennemeier y cols. [63] han señalado que los pacientes con lesiones talámicas pueden presentar dificultad para utilizar estrategias mnésicas, más que padecer un defecto de codificación de la información. Se ha propuesto que una interrupción entre el núcleo DM y el córtex prefrontal puede ser la responsable de la aparición de estos déficits.
Sin embargo, existen datos que ponen de manifiesto la aparición de un deterioro similar en la función ejecutiva después de infartos talámicos que no implican al núcleo DM. Se han descrito como lesiones en los núcleos intralaminares y partes adyacentes de los núcleos de la línea media pueden causar déficits en la función ejecutiva [52, 63].
Van der Werf y cols. [61] señalan que lesiones que impliquen a un único núcleo tálamico, por si mismas no son suficientes para que aparezca deterioro en la función ejecutiva,siendo necesaria la afectación de dos o más núcleos (DM, intralaminares y de la línea media).
REFERENCIAS
1. Sherman SM, Guillery RW. Exploring the Thalamus. San Diego: Academic Press; 2001 2. Carpenter MB. Neuroanatomía. Fundamentos. (4ªed.). Buenos Aires: Editorial Médica Panamericana; 1994 3. Ralston HJ. Tálamo. En Wong-Riley MMT, ed. Secretos de las Neurociencias. Mexico: McGraw-Hill; 2001. pp. 275-81. 4. Amaral DG. Organización funcional de la percepción y el movimiento. En Kandel ER, Schwartz JH, Jessell TM, eds. Principios de Neurociencia (4ªed.). Madrid: McGraw-Hill Interamericana 2001; pp. 337-48. 5. Elliot HC. Textbook of Neuroanatomy. Philadelphia: Lippincatt Co; 1969 6. Afifi AK, Bergman RA. Neuroanatomía Funcional. Texto y Atlas. México: McGraw-Hill Interamericana; 1999 7. Armengol JA. Centro y vías nerviosas I. Médula espinal, tronco del encéfalo y cerebelo. En Delgado JM, Ferrús A, Mora F, Rubia FJ, eds. Manual de Neurociencia. Madrid: Síntesis; 1998. pp. 359-392. 8. Morel A, Magnin M, Jeanmonod D. Multiarchitectonic and Stereotactic Atlas of the Human Thalamus. The Journal of Comparative Neurology 1997, 387: 588-630. 9. Hirai T, Jones, EG. A new parcellation of the human thalamus on the basis of histochemical staining. Brain Research Reviews 1989, 14: 1-34. 10. Martin JH. Neuroanatomía (2ªed.). Madrid: Prentice Hall; 1997 11. Steriade M, Deschenes M. The thalamus as a neuronal oscillator. Brain Research Reviews 1984, 8: 1-63. 12. Lorente de Nó R. Cerebral cortex: Architecture, intracortical connections, motor projections. En Fulton J, ed. Physiology of the Nervous System. Oxford: Oxford University Press; 1938. pp. 291-325. 13. Macchi, G. The intralaminar system revisited. En Minciacchi D, Molinari M, Macchi MG, Jones EG eds. Thalamic Networks for Relay and Modulation. Oxford: Pergamon Press; 1993. pp. 175-184. 14. Jones EG. Viewpoint: the core and matrix of thalamic organization. Neuroscience 1998; 85: 331-45. 15. Kuroda M, Yokofujita J, Murakami K. An ultrastructurd study of the neural circuit between the prefrontal cortex and the mediodorsal nucleus of the thalamus. Progress in Neurobiology 1998; 54(4): 417-58. 16. Bentivoglio M, Macchi C, Albanese A. The cortical projection of the thalamic intralaminar nuclei, as studied in cat and rat with multiple fluorescent retrograde tracing techique. Neurosci Lett 1981; 26: 5-10. 17. Stephens RB, Stilwell DL. Arteries and veins of the human brain. Springfiel: Charles C. Thomas; 1969. 18. Tatu L, Moulin Th, Bogousslavsky J, Duvernoy H. Arterial territories of the human brain. Neurology 1998; 50, 1699-1708. 19. Purves D, Augustine GJ, Fitzpatrick D, Katz LC, LaMantia AS, McNamara JO. Invitación a la Neurociencia. Madrid: Editorial Médica Panamerica; 2001 20. Ghika j, Bogousslavsky J, Maeder P y Regli F. The “Jerky distonic unsteady hand”: a delayed motor syndrome in posterior thalamic infarctions. Journal of Neurology 1994; 241: 537-42 21. Lee MS, Marsden CD. Movement disordres following lesions of the thalamus or subthalamic region. Movement Disorders 1994; 9(5): 493-507. 22. Lee MS, Kim YD, Yang JW, Lyoo CH, Oh SH, Kim HS. Clinical and anatomical factors associated with thalamic dyskinesias. Journal of the Neurological Sciences 2001; 182: 137-142. 23. Steriade M, Parent T, Hada A. Thalamic projections of nucleus reticularis thalami of cat: a study using retrograde transport of horseradish peroxidase and fluorescent tracers. J. Comp. Neurol 1984; 229: 531-47. 24. Royce GJ, Bromley S, Gracco C. Subcortical projections on the centromedian and parafascicular thalamic nuclei in the cat. J Comp Neurol 1991; 306: 129-55. 25. Parent A. Extrinsic connections of the basal ganglia. Trends in Neurosciences 1990; 13: 254-8. 26. Groenewegen HJ, Berendse HW. The specificity of the “nonspecific” midline and intralaminar thalamic nuclei. Trends in Neurosciences 1994; 17: 52-7. 27. Perea MV. Fundamentos de Neuropsicología. Psicobiología del Movimiento. Salamanca: Ediciones Universidad de Salamanca; 1989 28. Bhatnagar SC, Andy OJ. Neurociencia para el estudio de las alteraciones de la comunicación. Barcelona: Masson-Williams & Wilkins; 1997. 29. Gaffan D, Parker A. Mediodorsal thalamic function in scene memory in rhesus monkeys. Brain 2000; 123: 816-27. 30. Morison RS, Dempsey EW. A syudy of thalamocortical relations. Americam Journal of Physiology 1942; 135: 281-92. 31. Jasper HH. Diffuse projection systems: the integrative action of the thalamic reticular system. Electroencephalography and Clinical Neurophysiology 1949; 1: 405-19. 32. Moruzzi G, Magoun HW. Brain stem reticular formation and activation of the EEG. Electroencephalography and Clinical Neurophysiology 1949; 1: 455-73. 33. Steriade M, McCormick DA, Sejnowski TJ. Thalamocortical oscillations in the sleeping and aroused brain. Science 1993; 262: 679-85. 34. Destexhe A, Contreras D, Sejnowski TJ, Steriade M. A model os spindle rhythmicity in the isolated thalamic reticular nucleus. Journal of Neurophysiology 1994; 72: 803-18. 35. Hofle N, Paus T, Rutens D, Fiset P, Gotman J, Evans AC, Jones BE. Covariation of regional cerebral blood flow with delta and spindle activity during slow wave sleep in humans. Journal of Neurocience1997; 17: 4800-08. 36. Paus T, Zatorre RJ, Hofle N, Caramanos Z, Gotman J, Pretides M, Evans AC. Time-related changes in neural systems underlying attention and arousal during the performance of and auditory vigilance tesk. Journal of Cognitive Neuroscience 1997; 9(3): 392-408. 37. Kinomura S, Larsson J, Gulyás B, Roland PE. Activation by atttention of the human reticular formation and thalamic intralaminar nuclei. Sciencie1996; 271: 512-4. 38. Fiset P, Paus, T, Daloze Th, Plourde G, Meurte P, Bonhomme V, Hajj-Ali N, Backman SB, Evans AC. Brain mechanisms of Propofol-induced loss of consciousness in human: a Positron Emission Tomographic study. Journal of Neuroscience 1999; 19(13): 5506-13. 39. Chudasama Y, Muir JL. Visual attention in the rat: a role for the prelimbic cortex and thalamic nuclei?. Behavioral Neuroscience 2001; 115(2): 417-28. 40. Watson RT, Heilman KM. Thalamic neglect. Neurology 1979; 29: 690-4. 41. Graff-Radford NR, Damasio H, Yamada T, Eslinger PJ, Damasio AR. Nonhemorrhagic thalamic infarction. Brain; 1985; 108: 485-516. 42. Barrett AM, Schwartz RL, Crucian GP, Kim M, Heilman, KM. Attentional grasp in far extrapersonal space after thalamic infarction. Neuropsychologia 2000; 38: 778-84. 43. Penfield W, Roberts L. Speech and Brain Mechanisms. Princenton: Princenton University Press; 1959. 44. Mateer C. Asymmetric effects of thalamic stimulation on rate os speech. Neuropsychologica 1978; 16: 497-9. 45. Bhatnagar SC, Andy OJ. Allevation of acquired stuttering with human centremedian thalamic stimulation. Journal Neurology, Neurosurgery and Psychiatry 1989; 52: 1182-4. 46. Johnson M D, Ojemann GA. The role of the human thalamus in language and memory: Evidence from electrophysiological studies. Brain and Cognition 2000; 42(2): 218-30. 47. Crosson B. Subcortical functions in language in memory. New York: Guilford Press; 1992. 48. Ojemann GA. Asymmetric function of the thalamus in man. Annals of New York Academy of Science 1977; 299: 380-96. 49. Bentivoglio M, Aggleton JP, Mishkin M. The thalamus and memory formation. En Steriade M, Jones EG, McCormick DA, eds. Thalamus. Volume II. Experimental and clinical aspects. Oxford: Elservier Science Ltd; 1997; pp. 689-720. 50. Weiskrantz L. On issues and theories of the human amnesic syndrome. En Weinberger NM, McGaugh, JL, Lynch G, eds. Memory systems of the human brain: animal and human cognitive processes. New York: Guilford; 1985. 51. Guberman A, Stuss DT. The syndrome of bilateral paramedian thalamic infarction. Neurology 1983; 33: 540-6. 52. Graff-Radford NR, Tranel D, Van Hoese GW, Brandt JP. Diencephalic amnesia. Brain 1990; 113: 1-25. 53. Parkin AJ, Rees JE, Hunkin NM, Rose PE. Impairment of memory following discrete thalamic infarction. Neuropsychologia 1994; 32: 39-51. 54. Calabrese P, Haupts M, Markowitsch H, Gehlen W. Case Report: The cognitive-mnesic performance profile of a patient with bilateral asymmetrical thalamic infarction. International Journal of Neuroscience 1993; 71: 101-6. 55. Peru A, Fabbro F. Thalamic amnesia following venous infarction: evidence from a single case study. Brain and Cognition 1997; 33: 278-94. 56. Celerier A, Ognard R, Decorte L, Beracochea D. Deficits of spatial and non-spatial memory and of auditory fear conditioning following anterior thalamic lesions in mice: comparison with chronic alcohol consumption. The European Journal of Neuroscience 2000; 12(7): 2575-84. 57. Tranel D, Damasio AR. Neurobiological Foundations of Human Memory. En Baddeley AD, Wilson BA, Watts FN. eds. Handbook of Memory Disorders. Chichester: Wiley; 1995. pp. 27-47. 58. Victor M, Adams RD, Collins GH. The Wernicke-Korsakoff syndrome and related neurologic disorders due to alcoholism and malnutrition. (2nd ed.). Philadelphia: Davis; 1989. 59. Zola-Morgan S, Squire LR. Neuroanatomy of memory. Annual Review Neuroscience 1993; 16: 547-63. 60. Visser PJ, Krabbendam L, Verhey FRJ, Hofman PAM, Verhoeven WMA, Tuinier S, et al. Brain correlates of memory dysfunction in alcoholic Korsakoff’s syndrome. Journal of Neurology, Neurosurgery and Psychiatry 1999; 67(6): 774-8 61. Van der Werf YD, Witter MP, Uylings HBM, Jolles J. Neuropsychology of infarctions in the thalamus: a review. Neuropsychologia 2000; 38: 613-27. 62. Stuss DT, Guberman A, Nelson R, Larochelle S. The neuropsychology of paramediam thalamic infarction. Brain and Cognition 1988; 8: 348-78. 63. Mennemeier M, Fennell E, Valenstein E, Heilman KM. Contributions of the left intralaminar and medial thalamic nuclei to memory. Comparisons and report of a case. Archives of Neurology 1992; 49: 1050-8.
Share this:
Enriquerubio.net. El dolor persiste pero ya no me molesta
El lóbulo frontal es uno de los cuatro lóbulos de la corteza cerebral y constituye una región grande que está situada en la parte delantera del cerebro, justo detrás de la frente. Es el responsable de procesos cognitivos complejos, las llamadas funciones ejecutivas.
El lóbulo frontal se relaciona con el control de los impulsos, el juicio, la producción del lenguaje, la memoria funcional (de trabajo, de corto plazo), funciones motoras, comportamiento sexual, socialización y espontaneidad.
Su nombre, lóbulo frontal proviene del latín: lobus frontalis) es un área de la corteza cerebral de los vertebrados. Los lóbulos frontales son los más “modernos” filogenéticamente. Esto quiere decir que solamente los poseen de forma desarrollada los animales más complejos, Como los vertebrados y en especial los homínidos. En el lóbulo frontal se localizan funciones vitales para la supervivencia .
Las funciones del lóbulo frontal son múltiples y van desde lo puramente somático hasta espiritual solapando esta funciones entre sí
Esta estandarizado en tres áreas funcionales que tienen una representación anatómica. Teniendo en cuenta que el cerebro la relación forma conjunción es más idea de que real, pero que la necesitamos para entendernos.
Área motora o corteza motora: Proyecta el movimiento que tienen que realizar las extremidades y los movimientos faciales..
Área premotora o corteza premotora: Esta área proporciona el mecanismo para ejecutar los movimiento y selecciona los movimientos que van a ser ejecutados.
Area prefrontal o corteza prefrontal: Es la que controla los procesos cognitivos para que los movimientos, comportamientos y conductas que se vayan a realizar sean los apropiados al momento y lugar concreto.
Las alteraciones cognitivo-conductuales qué se producen por lesiones en esta área, se pueden estandarizar en:
Dificultad o incapacidad para formar estrategias y/o tomar decisiones correctas en la resolución de problemas.
Dificultad para anticipar, planificar, secuenciar y crear expectativas.
Disminución del pensamiento divergente, el cual está relacionado con la creatividad.
Disminución del habla. Pero no de pérdidas en la capacidad de comunicación, sino de aspectos relacionados con disminución del discurso y la espontaneidad oral, no variación en el tono de voz…
Disminución de la espontaneidad conductual general, como por ejemplo alegrarse tras una buena noticia, llorar por una mala…
Disminución de la flexibilidad conductual, lo cual altera la capacidad de buscar alternativas, cambios o estrategias en las situaciones del día a día. Y además genera rigidez mental produciendo perseverancia en pensamientos o ideas.
Dificultad para inhibir estímulos visuales externos, generando distracción y disminuyendo la capacidad de focalizar y controlar la mirada voluntaria.
Perdida de la memoria de trabajo, lo que dificulta mantener información en el tiempo y por lo tanto generar aprendizajes nuevos.
Disminución o pérdida de la conducta social, que es la que controla que tipo de comportamiento se debe realizar en cada contexto, y por lo tanto aparecerán conductas de desinhibición social como verborrea, irritabilidad, impulsividad, agresividad…
Modificación de la conducta sexual. Pueden aparecer conductas de desinhibición sexual o de indiferencia o disminución del interés sexual.
Disminución o pérdida de la capacidad de asociar acontecimientos personales a situaciones vividas.
A nivel cognitivo-conductual el lóbulo frontal esta encargado de controlar la mayoría de las capacidades relacionadas con la personalidad y las conductas de las persona
Relación de los lóbulos frontales, las funciones ejecutivas y las conductas
Los lóbulos prefrontales son el sustrato anatómico para las funciones ejecutivas, que son aquellas que nos permiten dirigir nuestra conducta hacia un fin y comprenden la atención, planificación, secuenciación y reorientación sobre nuestros actos.
Los lóbulos frontales tienen importantes conexiones con el resto del cerebro. Es el del director de orquesta; que dirige la información de todas las demás estructuras y las coordina para actuar sincrónicamente. Decía Goldberg, en El cerebro ejecutivo.
Los lóbulos frontales también están muy implicados en los componentes motivacionales (motivación) y conductuales (conducta) del sujeto;. por lo que si se produce un daño en esta estructura puede suceder que el sujeto mantenga una apariencia de normalidad al no existir déficits motrices, de habla, de memoria o incluso de razonamiento; existiendo sin embargo un importante déficit en las capacidades sociales y conductuales.
Este tipo de pacientes pueden ser por un lado apáticos, inhibidos… o por el contrario desinhibidos, impulsivos, poco considerados, socialmente incompetentes, egocéntricos, etcétera. Este tipo de déficits, al no ser tan evidentes como otros fueron los que llevaron durante mucho tiempo a los médicos a considerar a estos lóbulos como las estructuras «silentes»; es decir, sin función aparente. Solo recientemente se ha reconocido la importancia central del lóbulo frontal en nuestra actividad cognitiva.
El caso de Phineas Gage y Egas Moniz se consideran un prototipos de individuos que tras lesionarse ambos lóbulos frontales, cambiaron marcadamente su conducta.………………………………..
El primer caso en el que se describió un cambio de conducta debido a un daño frontal data de 1848, y fue el posteriormente famoso Phineas Gage, descrito por el doctor Harlow. Actualmente está considerado una de los casos clínicos clásicos dentro de la historia de la neurología y la neuropsicología cognitiva.
El reportaje clínico de Boston destaca la sorpresa del cuerpo médico por la supervivencia de Gage, que debería haber muerto instantáneamente; dice: «inmediatamente después del estallido Gage cayó de espaldas»; algo más tarde tuvo «movimientos convulsivos en las extremidades, pudiendo hablar a los pocos minutos»; los obreros (que le tenían mucho afecto) lo llevaron en brazos hasta la ruta, distante una veintena de metros, y lo subieron a una carreta que lo transportó un kilómetro, hasta el hotel de Joseph Adams; Gage estuvo sentado, muy erguido, todo el trayecto y después «se bajó de la carreta por sí mismo, ayudado por algunos de sus hombres».
John Harlow, uno de los médicos del pueblo. Mientras espera, supongo que dice, «pero señor Gage, ¿qué está pasando?» y una hora más tarde de la explosión, llega el doctor Edward Williams, colega más joven de Harlow. Años después describirá la escena como sigue: «Cuando llegué, Gage estaba sentado en una silla, en la galería del hotel de Adams, en Cavendish; me dijo ‘Doctor, aquí hay trabajo para usted’. Había visto la herida antes de bajar del coche, ya que las pulsaciones del cerebro eran patentes, pero sólo pude detallar su aspecto después del examen. La parte superior de la cabeza parecía un embudo invertido; en los bordes de la lesión, había pedazos de hueso; la apertura a través del cráneo e integumentos tenía unos tres centímetros de diámetro, y la herida parecía producida por un objeto en forma de cuña, que hubiera perforado de abajo hacia arriba. Mientras le examinaba la cabeza, Gage contaba a los mirones cómo había sucedido el accidente; se expresaba con tanto juicio que le hice directamente las preguntas del caso, en lugar de plantearlas a los testigos que lo acompañaban. Me relató, como haría muchas veces en años posteriores, algunos detalles del percance. Estoy en condiciones de afirmar que, en ningún momento, entonces o después, advertí en él algún síntoma de irracionalidad, excepto en una ocasión, a dos semanas del accidente, en que insistía en decirme John Kirwin, a pesar de lo cual me contestaba correctamente todas las preguntas». 3
Todo el episodio es sorprendente: sobrevivir a una explosión como ésa, y poder, a pesar de una enorme herida en el cráneo, hablar, caminar y ser coherente de inmediato, resulta caso increíble. Más asombroso aún es que Gage haya resistido la inevitable infección que se presentó en la herida,. Aunque en esos tiempos no hay antibióticos, el médico, con los productos químicos a su alcance, limpiará vigorosa y regularmente la llaga, y mantendrá al paciente en una posición inclinada para drenarla mejor. Gage tendrá un absceso —que Harlow quitará prestamente con su escalpelo— y fiebre alta, pero su contextura robusta y juvenil superará todos los inconvenientes.
El paciente será dado de alta en menos de dos meses. Sin embargo, ese increíble desenlace pierde relieve si se lo compara con el vuelco extraordinario que se producirá en la personalidad de Gage. Sus sueños, ambiciones, apetencias y desapetencias, están por cambiar. El cuerpo de Gage está vivo y bien, pero un nuevo espíritu lo anima.
Este caso ha sido extensamente investigado por el matrimonio Hanna y António Damásio, De forma que aunque los médicos que lo trataron en vida dieron abundantes anotaciones de su cambio de conducta, no es suficiente para localizar la lesión cerebral ya que al traumatismo que sufrió por la penetración de la de hierro en su cráneo, se siguió unos dias mas tarde de la supuración de la herida y es mas que probable que esta infección dañara además tejidos neurales adyacentes a la lesión y ésta fuera mas extensas que el propio lóbulo frontal. .
Lo más sorprendente de esta desagradable historia es la discrepancia de personalidad de Gage antes y después del accidente. Su normalidad se vio interrumpida por rasgos funestos que no desaparecieron jamás. Había sabido todo lo necesario para optar adecuadamente y ascender en la vida; tenía un marcado sentido de responsabilidad personal y social que se reflejaba en la forma como había logrado avanzar en su carrera profesional; era puntilloso en el trabajo y despertaba admiración en colegas y empleadores. Perfectamente adaptado a la sociedad, al parecer actuaba de manera escrupulosa y ética. Después del accidente se convirtió en un individuo irrespetuoso y amoral, cuyas decisiones no cuidaban sus intereses más elementales; se dio a inventar cuentos que «sólo nacían de su fantasía», según dice Harlow. El futuro no le interesaba y era absolutamente incapaz de preverlo
Si es válido en mi opinión la leucotomía prefrontal que practicó Almeida Lima en 1935, tras la insistencia de el neurólogo portugués Egas Moniz Y. La leucotomía prefrontal consistía en una ablación de los lóbulos prefrontales del cerebro y su objetivo era tratar trastornos mentales como la depresión. Egas Moniz afirmó tener buenos resultados popularizándose en todo el mundo y recibiendo éste el premio Nobel por ello en 1949. Sin embargo, la realidad era distinta y muchos de sus pacientes tuvieron fuertes cambios de personalidad que les incapacitaron para la vida en sociedad. Pese al atrevimiento de estos autores, es necesario recordar, la ausencia a lo del remedios para tratar a los enfermos psiquiatricos. No siempre la ciencias han sido virtuosas, ni la evolución tampoco lo ha sido, pero es lo que tenemos. Afortunadamente en nuestros días la investigación animal evita estos desastres. Y
Áreas del Lóbulo Frontal
Área precentral
Ubicada en la circunvolución precentral, por delante del Surco Central de Rolando y por detrás del Surco Precentral.
Se divide en:
Región posterior (área motora primaria o área 4 de Brodmann): Su función es llevar a cabo los movimientos individuales de diferentes partes del cuerpo. Recibe aferencias del tálamo, corteza sensitiva, área premotora, cerebelo y ganglios basales ya que esta área constituye la estación final para la conversión del diseño en la ejecución del movimiento.2
Región anterior (área motora secundaria, área premotora, o área 6 de Brodmann y partes de las áreas 8, 44 y 45): Almacena programas de actividad motora reunidos como resultado de la experiencia pasada. Participa en el control de movimientos posturales groseros mediante sus conexiones con los ganglios basales, además recibe aferencias de la corteza sensitiva y tálamo. Es la que programa la actividad del área motora primaria.
Esta área no produce perdida permanente del movimiento.3
Campo ocular frontal
Se encarga de los movimientos conjugados de los ojos, sobre todo los del lado opuesto. Controla los movimientos oculares voluntarios y es independiente de estímulos visuales.
Área motora del lenguaje o Área de Broca
Ubicada en la circunvolución frontal inferior, es importante en la formación de palabras, debido a sus conexiones con el área motora primaria. En la mayoría de las personas esta área es dominante en el hemisferio izquierdo, y la ablación del hemisferio no dominante no tiene efectos sobre el lenguaje, mientras que el daño del hemisferio dominante produce pérdida de la capacidad para producir la palabra, es decir una afasia de expresión, conocida como Afasia de Broca.
Corteza prefrontal
Se ubica por delante del área penetrante, región extensa que se conecta con un gran número de vías aferentes y eferentes. Se vincula con la personalidad del individuo y con la regulación de la profundidad de los sentimientos, así como en la determinación de la iniciativa y el juicio del individuo. También interviene en el proceso de atención.
Las lesiones de la corteza prefrontal se pueden presentar como un síndrome apático o pseudodepresivo, que se traduce en una reducción de la espontaneidad motora y verbal, pérdida de iniciativa, actividad motora y mental más lenta, indiferencia afectiva, escasa emotividad y menor interés sexual. (se relaciona con lesión de la región frontomedial).
Mientras otros presentan un síndrome desinhibido o pseudopsicopático, que se caracteriza por dificultad para reducir la velocidad de ciertas conductas, pérdida de autocrítica, conducta social inapropiada, indiferencia por los demás, y desinhibición o promiscuidad sexual (se relaciona con una lesión de la región frontobasal).
La corteza frontopolar es la parte de la corteza cerebral prefrontal, que ha evolucionado más recientemente y está relacionada con la planificación y el control de otras regiones cerebrales. Este corte realizado en la parte frontal del cerebro también revela otros elementos del cráneo, como los ojos, la cavidad nasal, los senos maxilares y la lengua.
Referencias
Snell, Richard S. (5ta edición). Neuroanatomía Clínica..
![VÍDEO] Encuentra un cerebro mientras camina por la playa](https://cronicaglobal.elespanol.com/uploads/s1/10/74/45/30/un-cerebro-humano-efe_10_670x355.jpeg)