.
Este articulo tiene por misión, criticar o por lo menos poner en duda una fisiología nerviosa que no es estable y que depende de lo interno y de lo interno y además cuando tienen proyecciones mentales, no tenemos reglas para medir.
Intentar expresarlos en términos aritméticos, tienen unas duda marcadas
Stephen W. Porges, Ph. D. Institute for Child Study, University of Maryland, College Park, M.D. 20742-
El autor de este trabajo considerado un puntal en el conocimiento del sistema vegetativo y sus reacciones describe minuciosamente el comportamiento de las motivaciones externas que conducen a una reacción patológica que se llama estrés .
Pero creo que no queda claro, que el estrés no es un estímulo externo es la respuesta a un estímulo externo y la respuesta patológica es la queja que manifiesta nuestra biología ante insultos externos.
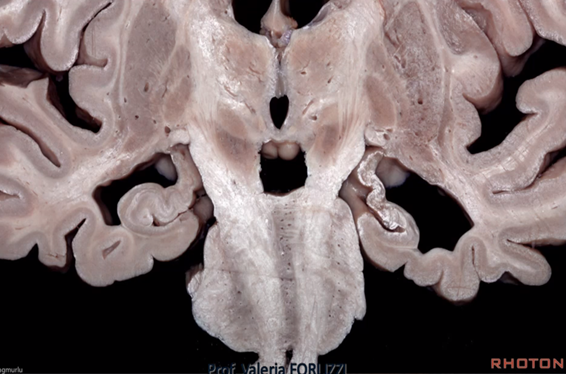
La organicidad de nuestro cerebro es tan compleja qué hace falta encajar de una manera acertada cómo funciona una complicada anatomía que es producto de la evolución de los seres vivos desde los animales unicelulares hasta el Homo sapiens.
Y la estructura que ha desarrollado nuestro cerebro es tan compleja porque es la suma de una serie de estructura que se han ido superponiendo con la evolución.
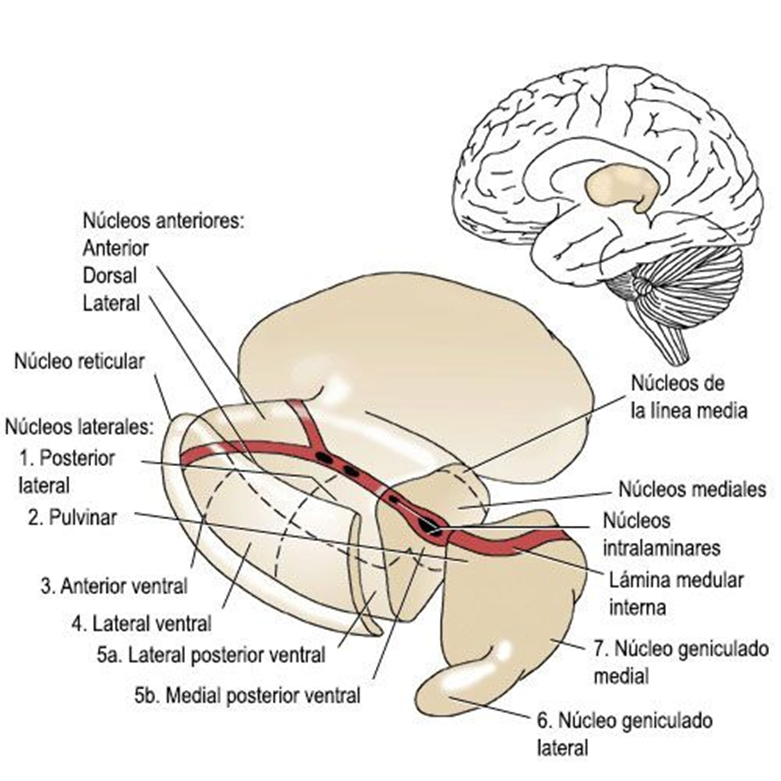
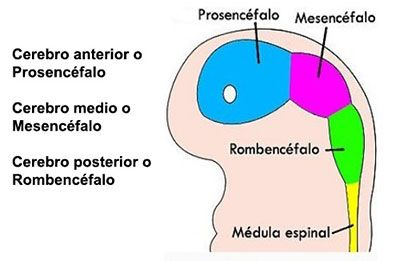
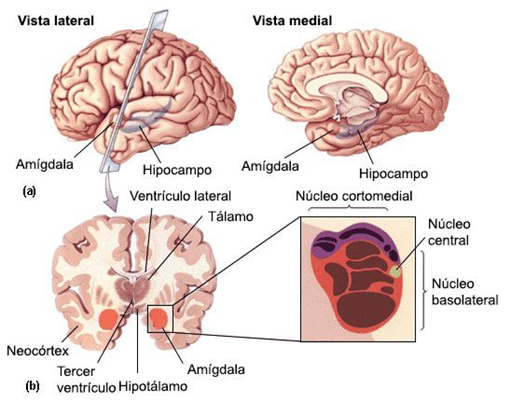
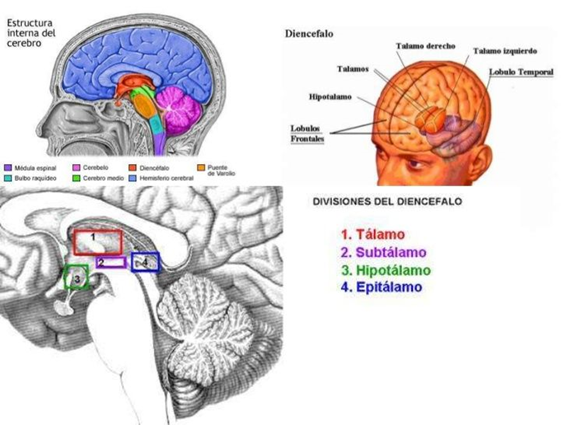
De forma que el cerebro Triuno de McClean, ofrece en mi opinión y de una manera gráfica cómo se van sumando cerebros desde los reptiles hasta el Homo sapiens y de una manera minuciosa el cerebro medio el Diencéfalo, el cerebro de las emociones tiene una especial complejidad porque la superposición evolutiva de estructuras redondeadas que forman el lóbulo límbico.
Esto está muy lejos de explicar la fisiología de esta estructura que hacer complicadísima anatómicamente lo es aún más cuando funciona. Y posiblemente porque el funcionamiento de las partes no es exclusiva sino que intervienen todas o varias y la respuesta será el resultado dependiendo de los múltiples estímulos que ocurren al mismo tiempo tanto externos como internos.
De forma que intentar darle una explicación al funcionamiento de esta estructura Triuna, donde determinadas actitudes son el resultado del funcionamiento de varias regiones al mismo tiempo no siempre correlacionadas producen una confusión al menos con el tipo de análisis que estamos utilizando..
El funcionamiento del sistema nervioso con los conocimientos que hoy tenemos es aleatorio.
El conocimiento parcial de los hechos desvirtua siempre los resultados.
La investigación sobre el estrés se ha enfocado, frecuentemente, en la descripción de eventos considerados como estresantes.
Por el contrario, en la pediatría, se asume que el estrés es un constructo fisiológico que es observado cuando la conducta se vuelve desorganizada y los procesos homeostáticos son interrumpidos. En los escenarios clínicos, esto se ha clasificado como inestabilidad fisiológica.
Existen amplias diferencias individuales en la vulnerabilidad a los mismos procedimientos médicos, las definiciones actuales del estrés enfatizan ya sea al tratamiento o a la respuesta al tratamiento y no al estatus neurofisiológico previo al tratamiento. Sin embargo, es este estado neurofisiológico el que puede identificar la vulnerabilidad del niño al estrés2.
Existe un método para valorar, de manera individual, tanto la respuesta al estrés como la vulnerabilidad al estrés. Este método monitorea el control neural del corazón vía el vago (Ej.; tono vagal) como un indicador de la homeostasis. El método permite la valoración de los efectos de procesos homeostáticos interrumpidos (Ej., estrés) y la vulnerabilidad de la homeostasis a ser alterada por varios tratamientos clínicos (Ej., vulnerabilidad al estrés).
ESTRÉS:
El sistema nervioso autónomo (SNA) regula la función homeostática, y está compuesto por dos subsistemas, el parasimpático (SNP) y el sistema nervioso simpático (SNS). El SNP y el SNS representan sistemas neurales que se originan en el tallo cerebral y contribuyen a la regulación de una variedad de órganos , incluyendo los ojos, glándulas lagrimales, glándulas salivales, glándulas sudoríparas, vasos sanguíneos, corazón, laringe, traquea, bronquios, pulmones, estómago, glándulas suprarrenales, riñones, páncreas, intestino, vejiga y genitales externos. En general, el SNP promueve funciones asociadas con un sistema de crecimiento y restauración. Por el contrario, el SNS promueve una respuesta metabólica incrementada para lidiar con desafíos externos al cuerpo.
En general, cuando un órgano visceral es inervado tanto por el SNS como por el SNP, los efectos son antagónicos. Por ejemplo, las neuronas del SNS dilatan la pupila, aceleran al corazón, inhiben los movimientos intestinales y contraen los esfínteres vesical y rectales. Las neuronas del SNP contraen la pupila, desaceleran al corazón, potencian el movimiento peristáltico y relajan los esfínteres rectal y vesical.
El SNP tiene fundamentalmente las actividades anabólicas que conciernen a la restauración y conservación de la energía y el reposo de los órganos vitales que sirven para la conservación del cuerpo; mediante la contracción de la pupila protegen a la retina del exceso de luz; mediante la desaceleración del ritmo cardíaco, dan al músculo cardíaco periodos más largos de descanso y vigorización; y al proveer al flujo de saliva y jugo gástrico y al suministrar el tono muscular necesario para la contracción del canal alimenticio, prueban ser fundamentalmente esenciales para el proceso de una digestión y absorción adecuadas mediante las cuales el material productor de energía es tomado y almacenado por el cuerpo. La gran labor de construir reservas y de fortalecer al cuerpo en tiempos de estrés y necesidad pertenece a la división craneal.
Se pueden ejecutar mediciones no sólo preventivas, sino incluso «proactivas», al identificar a los individuos que presentan inclinaciones a desarrollar (o no) ciertos estados disfuncionales dada la presencia de algunos factores de riesgo ya conocidos.
La estimulación del SNS prepara al individuo para la acción muscular intensa requerida para la protección y defensa como una respuesta a los desafíos externos. El SNS moviliza rápidamente las reservas existentes en el cuerpo. Los ojos se dilatan, la frecuencia y fuerza del corazón se incrementa con las contracciones, los vasos sanguíneos se encogen (vasoconstricción) y la presión sanguínea se incrementa. La sangre es drenada desde el depósito del intestino para alentar la disponibilidad y el transporte de sangre oxigenada a los músculos esqueléticos, pulmones, corazón y cerebro. La peristalsis y la secreción alimenticia son inhibidas y las contracciones de los esfínteres bloquean las salidas urinaria y rectal.
El SNS y el SNP son recíprocamente inervados y sus respuestas son coordinadas para proporcionar el estado interno adecuado para satisfacer las demandas de los cambios internos y externos. El SNP es modulado, primordialmente, por cambios internos en las vísceras. El SNS es activado, primordialmente, por impulsos exteroceptivos vía las fibras somáticas aferentes en respuesta a los cambios externos en el ambiente.
El SNP facilita la digestión y conserva la energía mediante la desaceleración de la frecuencia cardíaca. En la ausencia de desafíos externos (Ej., cambios en la temperatura ambiental, ruido, dolor, agentes pirogénicos, etc.), el SNP optimiza la función de las vísceras. Por otro lado, al aumentar la respuesta metabólica para lidiar directamente con desafíos externos, el SNS intenta optimizar la relación del organismo con el ambiente. En congruencia con esta descripción funcional del SNA, Gellhorn clasificó al SNP como un sistema trofotrópico y al SNS como un sistema ergotrópico.
Las primeras investigaciones anteriores definieron al SNA como un motor puramente visceral. Esta limitada definición no reconocía la contribución de los aferentes viscerales.
Las fibras aferentes acompañan a la mayoría de las fibras viscerales eferentes y forman la extremidad aferente de los reflejos viscerales. La visión contemporánea del SNA es que éste es un sistema complejo que contiene tanto eferentes periféricos como fibras aferentes así como estructuras neurales centrales.
Se deduce según Hess que la función del SNA refleja más un sistema nerviosos visceral que un sistema nervioso autónomo o vegetativo. Dado que el SNA es un sistema integrado por neuronas tanto periféricas como centrales, la medición de la actividad visceral periférica proporciona una ventana a las estructuras cerebrales que regulan la función y el estado visceral.
El SNA responde tanto a los estímulos externos como internos. A pesar de que el SNA es frecuentemente visto como un sistema motor que controla los órganos viscerales, la mayoría de las neuronas autónomas son aferentes. En el mantenimiento de las funciones corporales y en la reacción a las situaciones estresantes, los aferentes del SNA son cruciales. La retroalimentación aferente de los órganos viscerales frecuentemente regula el tono del SNP y tienen poco impacto en el tono del SNS. Por ejemplo, la distensión del estómago o la estimulación de los barorreceptores resultan en un incremento reflejo en el tono del SNP. La retroalimentación aferente de los órganos
sensoriales produce un perfil de respuesta diferente. Las respuestas autónomas a los estímulos
externos, incluyendo la nocicepción o atención, producen un decremento en el tono del SNP. Hay
un incremento complementario en el tono del SNS sólo si el estímulo es de alta intensidad y duración prolongada o está en condiciones de nocicepción o a estimulación intensa. En respuesta a las demandas metabólicas, las dos ramas del SNA frecuentemente funcionan sinergéticamente para maximizar la respuesta cardiovascular. Por ejemplo, durante el ejercicio hay un decremento progresivo del tono en el SNP y un incremento paralelo en el tono del SNS. Hay situaciones únicas en las que la respuesta autónoma está caracterizada ya sea por una activación dual o por una inhibición dual. Por ejemplo, en los humanos adultos, la respuesta sexual está caracterizada por una excitación dual.
El SNA no es solamente un sistema de respuesta que espera tranquilamente desafíos externos del ambiente. Por el contrario, el SNA está continuamente al servicio de las aferentes viscerales en un intento por mantener la homeostasis y promover la estabilidad fisiológica. Este proceso regulador es primordialmente mediado por el SNP. Desgraciadamente, hay estados de enfermedad que comprometen la función reguladora. Algunos estados de enfermedad (EG, hipertensión) están caracterizados por una depresión del tono del SNP con la compensación de una excitación en el SNS. Otros estados de enfermedad (Ej., diabetes) son caracterizados por una depresión en el tono del SNP sin evocar una excitación reciproca del SNS, mientras que otros estados de enfermedad pueden ser caracterizados por una inhibición dual.
El SNA está involucrado en la expresión fisiológica del estrés. Los cambios en la actividad del SNA que alteran los procesos homeostáticos parecen caracterizar el tema común asociado a la definición fisiológica del estrés. Una investigación más específica de la literatura sugiere que el SNP, virtualmente independiente del SNS, regula los proceso homeostáticos y sería, entonces, más sensible al estrés.
ESTRÉS Y HOMEOSTASIS: NUEVAS DEFINICIONES
Existen muchas definiciones del estrés. Dado que la mayoría de las definiciones están estructuradas en términos de las influencias causales (Ej., estímulo o contexto) y no en términos de variables comúnmente medidas en escenarios clínicos, dichas definiciones tienden a ser limitadas en su uso en el ambiente médico. Frecuentemente, las definiciones son circulares porque el estrés es definido tanto en términos del contexto (Ej., tratamiento médico) como de la respuesta (Ej., respuestas conductuales y fisiológicas). Por ejemplo, ¿es estresante un tratamiento médico porque provoca un aumento en la presión sanguínea y en la frecuencia cardíaca? O, ¿reflejan los incrementos en la presión sanguínea y en la frecuencia cardíaca un estrés independiente del tratamiento médico específico? O, ¿reflejan estrés las respuestas fisiológicas porque el clínico asume que existe estrés asociado al tratamiento médico? La definición es aún más confusa porque asumimos que existen diferencias individuales en la responsividad y vulnerabilidad al evento estresante. Por lo tanto, el tratamiento estresante puede no provocar una respuesta estresante en un paciente mientras que un tratamiento no estresante puede provocar una respuesta estresante en otro paciente.
Las definiciones actuales del estrés no son muy útiles en los escenarios clínicos. Aún si el estrés fuera definido operacionalmente al clasificar los estímulos estresantes como estresores y las respuestas conductales y fisiológicas al estresor como estrés, quedarían por lo menos dos problemas:
El estrés no debe ser conceptualizado simplemente en función del estresor y de la respuesta observada, sino también en cuanto al estado fisiológico de vulnerabilidad del paciente al momento del tratamiento. Nuevas definiciones del estrés y de la vulnerabilidad al estrés pueden ser derivadas y definidas operacionalmente basándose en la función del SNA. Las mediciones fisiológicas pueden ser valoradas objetivamente en escenarios clínicos mediante el monitoreo continuo del estrés y de la vulnerabilidad al estrés. Al desarrollar esta aproximación, se deben discutir dos áreas esenciales: (1) el fundamento para evaluar variables autónomas específicas como índices de estrés y (2) la tecnología de medición necesaria para medir los índices autónomos del estrés, continuamente, en un escenario clínico.
UNA DEFINICIÓN DE ESTRÉS DEL SNA: UNA FUNDAMENTACIÓN El SNA juega tanto con la distribución de los recursos para la atención de las necesidades de las vísceras internas como con la respuesta a desafíos externos. El sistema nervioso central media la distribución de recursos para lidiar con demandas internas y externas. Las percepciones y presuntas amenazas a la supervivencia, independientemente de las características físicas actuales de la estimulación, pueden promover una retirada masiva del tono del SNP y la activación recíproca del tono del SNS. El intercambio entre necesidades internas y externas puede ser usado para desarrollar definiciones del estrés y homeostasis. En base a este modelo, el estrés y la homeostasis son interdependientes. La homeostasis refleja la regulación de las vísceras internas y el estrés refleja la subordinación de necesidades internas en respuesta a necesidades externas. Por lo tanto, la medición del tono del SNP puede suministrar una variable indicadora para definir estrés y vulnerabilidad al estrés.
El concepto de homeostasis no es nuevo. Walter Cannon acuño el término y afirmó que “las reacciones fisiológicas coordinadas que mantienen la mayoría de los estados estables en el cuerpo son tan complejas y peculiares al organismo vivo que se ha sugerido el empleo de una designación específica para estos estados–homeostasis”. Las visiones de Cannon dependen del trabajo previo de Claude Bernard. El constructo “le milieu interieur” incluyó mecanismos fisiológicos responsables del mantenimiento de la constancia del ambiente interno. A medida que el trabajo de Bernard evolucionó, éste enfatizó la naturaleza dinámica y oscilatoria del sistema nervioso en el mantenimiento de un “internal milieu” dentro de un rango limitado. La homeostasis como un constructo nunca fue planeada para reflejar un estado estático. Más bien, definió la retroalimentación dinámica y el proceso de regulación necesario para el organismo vivo para mantener estados internos dentro de un rango funcional. Con el tiempo, ha perdido mucho de su rico significado y ha sido frecuentemente interpretado erróneamente para representar un nivel estático interno. Clínicamente, la “stasis” o falta de variabilidad endógena en sistemas periféricos neuralmente mediados, como la movilidad gástrica y la frecuencia cardiaca, es un signo de compromiso fisiológico severo3.
En el modelo propuesto, el SNP alimentan las necesidades viscerales (Ej., homeostasis) y el SNS responde a los desafíos externos. Por lo tanto, el estado del SNP se puede comparar con la homeostasis. Alternativamente, la retirada del tono en el SNP en respuesta a un desafío puede 3 Este señalamiento sugiere y reitera el hallazgo de Goldberger (1990) quien argumentó «que la reducción de la variabilidad precede u ocurre antes de la muerte cardíaca súbita»; en este sentido, la presencia de variabilidad fisiológica es un indicador general de salud y lo contrario de vulnerabilidad al estrés.
Definir el estrés, y el tono en el SNP previo al desafío puede representar vulnerabilidad fisiológica o vulnerabilidad al estrés. Con este modelo fisiológico, el estado del SNS no es una característica definitoria del estrés o de la vulnerabilidad al estrés; y la respuesta al estrés y la vulnerabilidad al estrés pueden ser clasificadas en la ausencia de cambios mayores en el tono en el SNS. Es importante notar que en muchas situaciones con niños sanos, la retirada transitoria del tono en el SNP tendrá como paralelo un incremento de la expresión del tono en el SNS. Por el contrario, niños severamente comprometidos pueden no exhibir reactividad en el SNS y el tono en el SNS puede ser bajo. También, estos niños tendrían un tono bajo en SNP, virtualmente ninguna reactividad en el SNP, y se valorarían clínicamente como crónicamente estresados y exhibiendo inestabilidad fisiológica.
Esta visión sugiere que la homeostasis puede definirse como el estado autónomo que alimenta las necesidades viscerales en ausencia de desafíos externos. Este estado estaría definido por un alto grado de tono el en SNP. Por lo tanto, el grado de estrés se puede cuantificar a un nivel fisiológico. Además, el estado autónomo crónico antes de un evento clínico podría clasificar la vulnerabilidad al estrés del paciente; los individuos que exhiban problemas de homeostasis tendrán la mayor vulnerabilidad al estrés.
LA VALORACIÓN DEL ESTRÉS: MONITOREO DEL TONO VAGAL Dada la anterior justificación, es necesario identificar y cuantificar una clasificación para la actividad del SNP. La medida más sencilla clasificada de la actividad del SNP se deriva del patrón de la frecuencia cardiaca. La amplitud de la arritmia sinusal respiratoria provee una clasificación válida y fácil para obtener el tono en el SNP vía el nervio vago cardiaco. En las siguientes secciones, esto se describirá como el tono vagal.
Con tecnología moderna es posible monitorear la influencia cambiante continua y en tiempo real del vago en el corazón y estimar cambios en el tono vagal general. Los procesos fisiológicos y conductuales dependen de la retroalimentación neural. La información es recibida en la periferia y transmitida al sistema nervioso central y los reflejos fisiológicos apropiados y las conductas manifiestas son emitidas. La retroalimentación se enlaza con muchos procesos homeostáticos y produce un patrón rítmico caracterizado por incrementos fásicos y decrementos en la respuesta neural eferente de órganos como el corazón. En muchos sistemas fisiológicos, el control neural eficiente se manifiesta como una variabilidad fisiología rítmica, y con parámetros normales, entre mayor sea la amplitud de oscilación, más sano es el individuo. Por lo tanto, la amplitud de los procesos fisiológicos rítmicos puede clasificar el estado del sistema nervioso del individuo y su capacidad para responder. En otras palabras, a mayor amplitud de la variabilidad fisiológica rítmica organizada, mayor el potencial de respuesta y el posible rango de conducta. Los individuos con variabilidad fisiológica atenuada podrían exhibir una falta de flexibilidad y conductual en respuesta a las demandas ambientales. Esto se observa en infantes muy enfermos. Por lo tanto, en función de las reacciones al estrés, uno podría esperar que en estos individuos faltara una capacidad autorreguladora para ajustarse rápidamente a los estímulos estresantes.
Investigaciones en las áreas de cardiología, gerontología, terapia física y diabetología demuestran que los déficits generales en el SNP están reflejados en el tono vagal cardiaco. Además, la estimulación de otras aferentes en el SNP resultará en un incremento reflejo en el tono vagal cardiaco. Dado que tono vagal refleja el estímulo general en el SNP a las vísceras y puede ser usado para monitorear el estrés y clasificar las diferencias en la vulnerabilidad al estrés.
Hemos desarrollado un índice del tono vagal. El índice del tono vagal es una medida de la modulación del sistema nervioso de la actividad de la frecuencia cardiaca vía el vago. El índice del tono vagal refleja influencias vagales eferentes rítmicas. Los patrones de frecuencia cardiaca, como los procesos conductuales, dependen del estado del sistema nervioso y de la calidad de la retroalimentación neural. El estrés resulta en una desorganización de la estructura rítmica tanto de la conducta como del estado autónomo. Por lo tanto, las mediciones del tono vagal cardiaco dan una importante visión del control central de los procesos autónomos necesarios para la conducta organizada. El tono vagal está reflejado en la amplitud del ritmo cardíaco asociado a la frecuencia de respiración espontanea. Este proceso rítmico, la arritmia sinusal respiratoria ha sido observada y estudiada por más de 100 años. Especulaciones en relación con los mecanismos neurales han sido reportadas desde 1910, y el tono vagal fue propuesto por Hering. Hering afirmó claramente que, “? se conoce con la respiración que una desaceleración demostrable de la frecuencia cardiaca? es indicativa de la función del vago”. Derivaciones de este método con las técnicas de respiración pausada son actualmente usadas para diagnosticar neuropatía periférica en pacientes diabéticos. La investigación contemporánea ha dado evidencia empírica de que la amplitud de la arritmia sinusal respiratoria traza exactamente la influencia eferente del nervio vago en el corazón. En base a estudios electrofisiológicos, se ha propuesto que el impulso respiratorio central da entrada al núcleo fuente de las fibras vagales cardioinhibitorias. Si el tono vagal es un índice sensible al estatus funcional del sistema nervioso, entonces predeciríamos que los individuos que los individuos con un mayor tono vagal exhibirían un rango mayor de conductas competentes. También esperaríamos que las condiciones que comprometen al sistema nervioso central (Ej., complicaciones médicas, anestesia y enfermedad) resultarían en una atenuación del tono vagal.
Al construir un modelo que relaciona el tono vagal con el estrés, describiremos primero una característica metafórica global de los procesos homeostáticos y luego más específicamente, de las vías relacionadas con el vago. La frecuencia cardiaca en un humano sano no es estable. El patrón de frecuencia cardiaca refleja la retroalimentación continua entre el sistema nervioso central y los receptores autónomos periféricos. La fuente principal de variabilidad de la frecuencia cardiaca es mediada por un incremento fásico y decrementos en la respuesta neural eferente vía el vago al corazón. Bajo la mayoría de las condiciones, como otras medidas de control homeostático, entre más grande es el rango de incrementos y decrementos fásicos, “más sano4” es el individuo. Por ejemplo, en los recién nacidos con alto riesgo, hay una atenuación del rango de funciones homeostáticas. En forma paralela a este proceso, hay también una reducción en el tono vagal.
La variabilidad en la frecuencia cardiaca es un indicador de la eficiencia de los mecanismos de retroalimentación neural y puede clasificar a los estados de salud o a la capacidad del individuo para organizar fuentes fisiológicas para responder apropiadamente. Por lo tanto, entre mayor sea la variabilidad fisiológica organizada, mayor será el rango de conducta. La variabilidad organizada se cuando se habla de un «mayor rango de conductas» se hace referencia también a los estilos de enfrontamiento que resultan más adaptativos por su variedad y no por la frecuencia en que son utilizados.
Este tipo de ciclos pueden observarse durante la obtención del «Perfil Psicofisiológico del Estrés» incluso para la retroalimentación dinámica, por lo que ésta está caracterizada por desviaciones rítmicas a un nivel constante. Un sistema de retroalimentación óptima permite grandes desviaciones de la media con una retroalimentación negativa ocurriendo a intervalos constantes. La regulación neural del corazón en individuos sanos es similar al sistema de retroalimentación óptimo, descrito anteriormente. En el control neural del corazón, el vago sirve como un mecanismo de retroalimentación primaria negativa. Por lo tanto, los estados de variabilidad de la frecuencia cardíaca atenuada tendrían influencias vagales atenuadas en el corazón.
Regresando a nuestra metáfora, los estados caracterizados por influencias vagales atenuadas deberían ser paralelos a la flexibilidad conductual reducida en respuesta a las demandas ambientales.
VULNERABILIDAD AL ESTRÉS EN EL RECIÉN NACIDO: UN EJEMPLO DE MONITOREO DEL TONO VAGAL
En las graficas que presenta del monitoreo de niños despiertos y dormidos La aclaración del concepto de tono vagal puede obtenerse mediante la observación de las diferencias entre el panel superior e inferior. Se puede notar que, incluso durante el sueño, la frecuencia cardiaca no es constante. A pesar de que los niveles de frecuencia cardiaca son similares para los dos recién nacidos, el más sano presenta mayor variabilidad entre latido y latido con relación al de alto riesgo. En una mejor inspección de los dos patrones de latidos se puede identificar una diferencia impresionante en los cambios rápidos de la frecuencia cardiaca que ocurren cada 1 a 3 segundos. Estas oscilaciones están asociadas a la respiración y reflejan el tono vagal cardiaco. El tono vagal cardiaco refleja la amplitud de estas rápidas oscilaciones y se reporta en unidades logarítmicas naturales.
Los sujetos en la muestra fueron 125 recién nacidos de gestación completa y 112 recién nacidos instalados en la unidad de terapia intensiva. Los recién nacidos de gestación completa fueron todos residentes de una guardería normal y fueron examinados durante el segundo día posterior al parto.Los valores del tono vagal de los prematuros fueron hallados en el primer registro hecho con los recién nacidos, sin usar el ventilador y dejando que respiraran el aire de la habitación.
El tono vagal fue valorado durante el sueño cuando los prematuros tenían entre 35 y 37 semanas corregidas de gestación. Los recién nacidos de alto riesgo, como grupo, tiene un tomo vagal significativamente bajo en comparación a los recién nacidos normales Dado que la frecuencia respiratoria puede influenciar la estimación del tono vagal, la respiración se monitoreó en un subgrupo de 47 recién nacidos normales y 62 recién nacidos prematuros. La respiración fue significativamente más rápida para los recién nacidos prematuros (F[1, 107] = 23.5, P .0001). De cualquier manera, aún cuando esta influencia significativa fue removida del análisis de varianza, todavía había una diferencia altamente significativa entre los dos grupos del tono vagal (F [1, 107] = 82.2, P .00001). La clasificación de grupo (recién nacidos normales vs. recién nacidos prematuros) explican el 53.1% de la varianza en el tono vagal. Cuando la influencia de la respiración fue removida, la clasificación de grupo explicó el 43.7% de la varianza en el tono vagal. Investigaciones recientes en nuestro laboratorio sugieren tan sólo un ligero incremento de maduracionj y una gran estabilidad de estas estimaciones bajo condiciones estándares de muestreo durante la residencia del grupo de prematuros. Por ejemplo, una muestra de 16 recién nacidos prematuros fue examinada durante el sueño en 5 días aislados, empezando por lo menos un día después del parto. A pesar de que hubo una relación significativa entre la severidad de la condición clínica y el tono vagal (Ej, recién nacidos más sanos y un tono vagal más elevado), la correlación promedio entre los valores del tono vagal fue cercana al .9. Lo anterior proporciona un ejemplo de cómo el monitoreo del tono vagal en los bebés prematuros puede suministrar una clasificación sensible a la vulnerabilidad al estrés. Datos de nuestro laboratorio y otros laboratorios demuestran que los residentes más comprometidos tienen un tono vagal más bajo. En consistencia con el modelo de estrés y vulnerabilidad de estrés anterior, los recién nacidos prematuros tienen un tono en el SNP limitado para regular su estado interno y son confrontados, simultáneamente, con demandas ambientales, que incluye la necesidad de termorregular y lidiar con los estímulos sensoriales e incluyendo los procedimientos médicos asumidos como dolorosos.El sistema vagal responde a las necesidades cambiantes del organismo. Con frecuencia, el sistema vagal reaccionará selectivamente incrementando o decrementando su influencia en la periferia. Esto puede ser observado como un abandono del tono vagal de incrementar la frecuencia cardiaca para atender las demandas metabólicas o un incremento en el tono vagal para regular los polipéptidos digestivos y la movilidad gástrica. El éxito adaptativo del infante no está basado meramente en el nivel tónico del SNA, sino en la habilidad del SNA de responder apropiadamente ante los desafíos ambientales e intervalares. Por ejemplo, durante las manipulaciones médicas ambientales como la circuncisión, el tono vagal es suprimido. Por el contrario, durante la alimentación GAVAGE de los recién nacidos prematuros, el tono vagal es incrementado. Si los infantes exhibieron un incremento en el tono vagal durante la alimentación GAVAGE y una depresión del tono vagal inferior a los niveles PREGAVAGE después de la alimentación, éstos fueron dados de alta aproximadamente dos semanas antes que los infantes que no exhibieron esta respuesta. Estos efectos fueron independientes del peso, tiempo de gestación y otros factores al momento del nacimiento. A pesar de que el tono vagal PREGAVAGE no fue relacionado al patrón de respuesta o al haber sido dados de alta, si predijo trayectorias de aumento de peso. Por lo tanto, la reactividad vagal proporciona otra dimensión relacionada al riesgo clínico.
Mediciones del tono vagal proporcionan una importante visión a la modulación central de la función autónoma. Mediciones del tono vagal durante el sueño o bajo condiciones no desafiantes proporcionan una clasificación de la retroalimentación homeostática normal, mientras que, mediciones hechas durante desafíos sensoriales o cognoscitivos proporcionan una indicación de un funcionamiento adaptativo. Por lo tanto, los niveles bajos en el tono vagal son asociados a poblaciones de alto riesgo, mientras que la responsividad vagal típica, incluso en infantes con niveles basales normales de tono vagal, parece identificar a un subgrupo de infantes que presentan problemas conductuales y regulatorios.
CAMBIOS EN EL TONO VAGAL DURANTE EL ESTRÉS: EL COSTO DE HACER NEGOCIOS
El SNA tiene muchas responsabilidades fisiológicas. Debe regular la presión sanguínea para asegurarse de que llegue al cerebro suficiente sangre. También monitorea a los gases sanguíneos. Si hay cambios en el oxígeno o el bióxido de carbono, inmediatamente se implementan cambios en los parámetros cardiopulmonares mediante la modulación neural directa del corazón, del tono vasomotor y de los pulmones. Mientras estos procesos cardiopulmonares están iendo regulados, el SNA está también controlando la digestión y el metabolismo. Las acciones del SNA están relacionadas con el mantenimientos de la vida: funciones ergotrópicas (Ej., trabajo) y trofotrópicas (Ej., crecimiento).
El sistema vagal es crítico para los procesos ergotrópicos y trofotrópicos. Los incrementos en el tono vagal no sólo resultan en incrementos de la respuesta metabólica, sino que modulan los polipéptidos digestivos y la movilidad gástrica. El vago es también crítico en la facilitación de los procesos trofotrópicos. El vago puede tener influencias inhibitorias directas en la excitación simpática del miocardio. Por otro lado, el sistema límbico, asumido por los psicofisiólogos como modulador del impulso autónomo a través de la excitación simpática, tiene influencias inhibitorias directas en las células de origen del vago. Las regiones del tallo cerebral que controlan las eferentes del vago actúan para expresar un mayor tono vagal, manteniendo los estados trofotrópicos, o para permitir la retirada del tono vagal y facilitar la movilización inmediata del organismo.
La investigación con otras especies demuestra que el tono vagal se incrementa durante el desarrollo.
En forma paralela a este incremento en el tono vagal, están los incrementos en las conductas autorreguladoras y exploratorias. En la investigación con infantes se ha reportado que un tono vagal alto está asociado a la memoria visual de reconocimiento. Un capítulo general resume la investigación con el tono vagal y su afectación.
DiPietro y Porges también evaluaron a los recién nacidos en relación al tono vagal y la reactividad conductual en la alimentación GAVAGE. En este estudio, las diferencias individuales en le tono vagal correlacionaron significativamente con la reactividad conductual ante el método GAVAGE de alimentación.
Similarmente, Huffman et al. observaron que los infantes de tres meses de edad con un alto tono vagal se habituaron más rápidamente a los estímulos habituales novedosos y exhibieron una mayor atención sostenida. Por ejemplo, en un estudio que evaluó los efectos de la ATROPINE SULFATE en el desempeño sensoriomotor, reportamos no sólo una depresión del tono vagal dependiente de la dosis, sino también decrementos dependientes de dosis en el desempeño. La anestesia inhalada deprimió el tono vagal y mientras los pacientes recuperaban la consciencia, hubo un incremento paralelo del tono vagal.
Con el desarrollo del índice del tono vagal hemos sido capaces de obtener una valoración más exacta de los mecanismos vagales y de la relación entre el tono vagal y la reactividad autonómica. Los estudios que usan la clasificación del tono vagal apoyan la hipótesis de que éste puede clasificar el estrés y la vulnerabilidad al estrés. Porter et al. demostraron, en una muestra de recién nacidos, una retirada masiva del tono vagal durante la circuncisión. Además, las diferencias individuales en el tono vagal correlacionaron con la reactividad de la frecuencia cardiaca a la circuncisión. Los recién nacidos con un tono vagal más alto exhibieron no sólo una mayor aceleración del ritmo cardíaco sino también frecuencias fundamentales de llanto bajas. Porter y Porges también demostraron, en infantes prematuros, que las diferencias individuales en el tono vagal estuvieron relacionadas a las respuestas de la frecuencia cardiaca durante procedimientos de perforación lumbar.
CONCLUSIÓN
Existe una base fisiológica para definir al estrés y a la vulnerabilidad al estrés. Los conceptos de estrés y homeostasis son interdependientes y manifiestos en la actividad del SNP. En comparación con los modelos tradicionales del estrés, el SNP se propone como un modulador de la vulnerabilidad al estrés y de la reactividad. El modelo propuesto sugiere que el monitoreo proporciona una ventana que permite la valoración del estrés.
Se presenta una justificación fisiológica para usar el SNP, en vez de o junto con la actividad del SNS, en la descripción de la vulnerabilidad al estrés y de la reactividad al estrés. La cuantificación del tono vagal cardiaco de las oscilaciones de la frecuencia cardíaca espontánea, se describe como un método para evaluar los estados cambiantes del SNP y proporciona además un instrumento estándar con parámetros estadísticos que son comparables entre pacientes y a través del ciclo vital.
El modelo no depende de etapas de desarrollo motor o cognoscitivo, por lo que su uso es práctico incluso con recién nacidos. Este método no invasivo permitirá la valoración del impacto estresante de varios tratamientos clínicos en el infante joven así como la identificación de individuos con vulnerabilidad al estrés.
El tono vagal es propuesto como un índice novedoso de la vulnerabilidad y reactividad al estrés, con aplicaciones en todas las ramas de la medicina y con un valor particular en la pediatría.
REFERENCIAS
- Cannon WB. Bodily Changes in Pain Hunger, Fear and Rage. An Account of Recent
Researches Into the Function of Emotional Excitement . New York, NY:D. Appleton; 1929.
- Gellhorn E. Principles of Autonomic-Somatic Integrations: Physiological Basis and
Psychological and Clinical Implications. Minneapolis, MN: University of Minnesota Press;
1967.
- Langley JN. The Autonomic Nervous System . London. UK: Heffer and Sons; 1921:1
- Hess WR. Diencephalon, Autonomic and Extrapyramidal Functions. New York, NY: Grune
and Stratton; 1954.
- Cannon WB. Organization for Physiological Homeostasis. Physiol Rev. 1929;9:399-431.
- Bernard C. Lessons on the phenomena of life common to animals and vegetables. Second
lecture: the three forms of life. 1978-1979. Reprinted in Langley L, ed. Homeostasis: Origins
of the Concept . Stroudsburg, PA:Dowden Hutchinson & Ross; 1973-129-151.
- Porges SW. Respiratory sinus arrhythmia: physiological basis, quantitative methods, and clinical
implications. In: Grossman, P, Janssen F, Vaitl D, eds. Cardiorespiratory and Cardiosomatic
Psychophysiology. New York, NY: Plenum:1986:101-115.
- Porges SW. Method and apparatus for evaluating rhythmic oscillations in aperiodic
physiological response systems. US patent no. 4520944; 1985.
- Cottingham JT, Porges SW, Lynon T. Soft tissue mobilization (Rolfing pelvic lift) and
associated changes in parasympathetic tone in two age groups. Phys Ther. 1988;68: 352-356.
- DiPietro JA, Porges SW. Vagal responsiveness to gavage feeding as an index of preterm status.
Pediatr Res. 1991;29:231-236.
- Goldberger AL, and Ridney D. R. (1990). Sudden death is not chaos. En: The ubiquity of chaos.
- Krasner (ed.), pp. 23-34.
- Hering HE. A functional test of heart vagi in man. Menschen München Med Wchnschy,
1910;57:1931-1933.
- Jordan D, Khalid MEM, Schneiderman N, et al. The location and properties of preganglionic
vagal cardiomotor neurones in the rabbit. Pfluegers Arch. 1982;395:244.
- Porges SW, McCabe PM, Yongue BG, Respiratory-heart rate interactions: psychophysiological
implications for pathophysiology and behavior. IN: Caccippo J, Petty R, eds. Perspectives in
cardiovascular psychophysiology. New York, NY: Guilford; 1982: 223-264.
- Fox NA, Porges SW. The relationship between developmental outcome and neonatal heart
period patterns. Child Dev, 1985;56:28-37.
13
- Porges SW, Neonatal vagal tone: diagnostic and prognostic: Vietze PN, Vaughn HG, eds. Early
Identification of Infants With Developmental Disabilities. Philadelphia, PA: Grune and Stratton;
1988:147-159.
- Porter FL, Porges SW, Marshall RE. Newborn pain cries and vagal tone:parallel changes in
response to circumcision. Child Dev. 1988;59:495-505.
- DeGangi GA, DiPietro JA, Greenspan SI, Porges SW. Psychophysiological characteristics of
the regulatory disordered infant. Infant Behav. 1991;14:37-50.
- Uvnas-Moberg K Gastrointestinal hormones in mother and infant. Acta Paediatr Scand Suppl.
1989;351:88-93.
- Levy MN. Parasympathetic control of the heart. In: Randall WC, ed.. Neural Regulation of the
Hearth. New York, NY: Oxford University Press; 1977.
- Schwaber JS, Kapp BS, Higgins G. The origin and the extent of direct amygdala projections to
the region of the dorsal motor nucleus of the vagus and the nucleus of the solitary tract. Neurosci
Lett. 1988;20:15-20.
- Larson SK, Porges SW. The ontogeny of heart period patterning in the rat. Dev. Psychobiol.
1982;15:519-528.
- Linnemeryer SA, Porges SW. Recognition memory and cardiac vagal tone in 6-moth-old infants.
Infant Behav. 14986;9:43-56.
- Richards JE. Respiratory sinus arrhythmia predicts heart rate and visual responses during visual
attention in 14-and 20-week-old infants. Psychophysiology. 1985;22:101-109.
- Porges SW. Vagal tone: a mediator of affect. In: Garber JA, Dodge KA, eds. The Development
of Affect Regulation and Dysregulation. New York, NY: Cambridge University Press; 1990.
- Huffman LC, Bryan YE, Pedersen FA, Porges SW. Infant temperament: relationships with heart
rate variability. Presented at the Annual Meeting of the Society for Behavioral Pediatrics.
Washington, DC, May, 1988.
- Dellinger JA, Taylor HL, Porges SW. Atropine sulfate effects on aviator performance and on
respiratory-heart rate period interactions. A. Space Evirnon Med. 1987; 58:333.
- Donchin, Y, Feld JM, Porges SW. The measurement of respiratory sinus arrythmia during
recovery from isoflurane-nitrous-nitrous oxide anesthesia. Anesth Analg. 1985;64:811.
- Porter FL, Porges SW. Neonatal cardiac responses to lumbar punctures. Infant Behav.
1988;11:261. Abstrac.
Traducción y comentarios de: B. Domínguez. Fac. Psic. UNAM, CNDH y Clínica del Dolor, Hospital Gral. De
México, S.S. E-mail: <benajmin@servidor.unam.mx>