EL PROPIO CUERPO ‘FABRICA’ EL TRATAMIENTO
Premio Fronteras a Karikó, Langer y Weissman por crear dos tecnologías que unidas han impulsado las terapias de ARN mensajero, abriendo la puerta al desarrollo de vacunas y tratamientos contra múltiples enfermedades
Entrevista a Katalin Karikó

Fotografía: De izquierda a derecha: Drew Weissman, Katalin Karikó y Robert Langer. Vídeo: Lectura del acta del jurado por su presidenta, la profesora Angelika Schnieke.
El Premio Fundación BBVA Fronteras del Conocimiento en Biología y Biomedicina ha sido concedido en su decimocuarta edición a Katalin Karikó, Robert Langer y Drew Weissman “por sus contribuciones a las terapias de ARN mensajero (ARNm) y a la tecnología de transferencia que permite a nuestras propias células producir proteínas para la prevención y el tratamiento de enfermedades”, señala el acta del jurado.
26 enero, 2022
La aplicación más sobresaliente derivada hasta ahora del trabajo de los galardonados es “el desarrollo oportuno y rápido de vacunas contra el SARS-CoV-2”, que han demostrado proporcionar “una protección eficaz contra la Covid-19 grave”. El jurado destaca que las vacunas que están conteniendo la pandemia son solo el principio de una tecnología “llamada a extenderse a otras áreas terapéuticas, como las enfermedades autoinmunes, el cáncer, los trastornos neurodegenerativos, las deficiencias enzimáticas y otras infecciones víricas”, señala el acta.
“Este premio reconoce a los creadores de las dos tecnologías que, unidas, no solo han hecho posible las vacunas contra el Covid-19, sino que abren todo un abanico de posibilidades terapéuticas en áreas muy diversas para el futuro. Las vacunas han sido el primer ejemplo del potencial de la unión de estas dos tecnologías, pero ya se está investigando y hay ensayos clínicos sobre su uso contra otras enfermedades”, explica Óscar Marín, director del Centro de Trastornos del Neurodesarrollo en King’s College London (Reino Unido) y secretario del jurado.
Karikó y Weissman, bioquímica e inmunólogo respectivamente, y Langer, ingeniero químico, son autores de avances cruciales en la cadena de hallazgos científicos que han convertido en realidad las llamadas terapias de ARN mensajero, una tecnología que logra que sean las propias células del cuerpo las que producen las moléculas con capacidad terapéutica.
Las nominaciones de Karikó y Weissman al Premio Fronteras del Conocimiento fueron hechas por Isabel Varela, presidenta de la Sociedad Española de Bioquímica y Biología Molecular (SEBBM), Larry Jameson, decano de la Escuela Perelman de Medicina y vicepresidente de la Universidad de Pensilvania, Eric Topol, vicepresidente del Scripps Research Institute, y Elias Zerhouni, catedrático emérito de la Universidad Johns Hopkins. La nominación de Langer fue realizada por Antonio López Díaz, Rector Magnífico de la Universidade de Santiago de Compostela, y por María José Alonso, catedrática de Farmacia y Tecnología Farmacéutica de la misma universidad.
Temporalmente, la primera contribución es la de Langer, catedrático del Instituto Tecnológico de Massachusetts (MIT, EE.UU.). En los años setenta del pasado siglo, Langer publicó en la revista Nature el primer trabajo que demostraba que era posible encapsular en nanopartículas moléculas de ácidos nucleicos −como el ARN, siglas de ácido ribonucleico−, y transferirlas al interior del cuerpo. Abría la puerta así a “empaquetar las macromoléculas terapéuticas, incluido el ARNm, de forma que puedan ser transferidas a las células, y que la propia maquinaria de traducción celular sintetice la proteína/antígeno”, explica el acta del jurado.
La aportación de Karikó y Weissman, ambos catedráticos en la Universidad de Pensilvania (EEUU), llegó ya entrado el nuevo siglo. Como recoge el acta, “juntos desarrollaron métodos de modificación del ARNm para evitar su destrucción por parte del sistema inmunitario humano”, una vez introducido en el organismo. Fue un avance clave.
“Karikó y Weissman descubrieron cómo modificar las moléculas de ARNm para hacerlas susceptibles de ser utilizadas como agente terapéutico, y Langer ideó el vehículo seguro, la tecnología de encapsulación que permite introducir el ARNm dentro del cuerpo”, explica Oscar Marín. “Los dos avances son imprescindibles”.
La molécula que transporta la información para sintetizar proteínas
Entrevista a Drew Weissman
El ADN y el ARN son las moléculas –químicamente son ácidos nucleicos– que contienen la información necesaria para que todo organismo vivo fabrique sus proteínas. El ADN de cada ser vivo es único y está presente en todas sus células. La función del ARN −en términos muy básicos− es copiar la información del ADN y transportarla hasta la maquinaria de la célula que se ocupa de fabricar las proteínas. El concepto de terapia de ARN parte de la base de que es posible diseñar ARN ‘a la carta’ en el laboratorio, de forma que contenga la información necesaria para fabricar cualquier proteína, ya sea un compuesto terapéutico o, como en las vacunas frente a la Covid-19, un fragmento de un virus. Una vez dentro de la célula, ese ARN sintético será leído por la maquinaria celular, que empezará a producir las proteínas deseadas.
Las vacunas de ARNm contra la Covid-19 contienen ARN con instrucciones para fabricar la proteína S del coronavirus SARS-CoV-2, que es la que actúa como llave para entrar en las células humanas. De esta forma, cuando la vacuna es inyectada, los macrófagos —un tipo de células defensivas del sistema inmune— próximos al lugar del pinchazo ingieren el ARN envuelto en grasa; estas células empezarán a producir la proteína S del virus y colocarla en su membrana externa, para exhibirla al exterior. Esto induce en el organismo una respuesta defensiva como la que se generaría para protegernos de una infección natural del SARS-CoV-2.
Estas vacunas se producen más rápido que las tradicionales, y pueden adaptarse más fácilmente a las mutaciones del virus. También son vacunas teóricamente más seguras, puesto que no interviene en el proceso ningún virus vivo, y ningún material genético entra en el núcleo de la célula humana.
El principio de una revolución biomédica con múltiples aplicaciones
Entrevista a Robert Langer
Tras conocer el fallo del jurado, Katalin Karikó ha explicado cómo se siente ahora que el éxito de las vacunas ha colocado su trabajo en un lugar central de la ciencia: “Durante 40 años no solo no recibí ningún premio, sino que no recibí ningún apoyo económico para mi investigación, así que este reconocimiento es un gran honor. Quiero aprovechar que estoy bajo los focos de los medios para animar a los jóvenes a dedicarse a la ciencia, porque es apasionante”.
Karikó empezó a trabajar con ARN sintético a finales de los setenta en el Centro de Investigaciones Biológicas de Szeged, en Hungría, su país natal. En 1985 emigró con su familia a Estados Unidos. Ya en la Universidad de Pensilvania la investigadora siguió trabajando en la tecnología del ARNm, una línea de investigación por la que pocos apostaban. El punto de inflexión fue el inicio de la colaboración con el inmunólogo Drew Weissman, de la misma universidad. En 2005 Karikó y Weissman lograron su primer gran avance: descubrir cómo modificar el ARN de forma que el sistema inmunitario humano no logre detectarlo.
Como ha explicado el propio Weissman tras conocer el fallo, “nuestra hipótesis central cuando empezamos este trabajo fue que el ARN sería un sistema mejor para transferir proteínas al organismo, porque convertiría al propio cuerpo receptor en la fábrica que produce la terapia. El problema que nos encontramos es que el ARN era enormemente inflamatorio, y el animal al que se lo inyectábamos se ponía enfermo, así que Katalin y yo estuvimos muchos años intentando averiguar la causa de este problema, y fue así como conseguimos nuestro hallazgo principal: un método para evitar la reacción inflamatoria del ARN. Esto además tuvo el efecto de incrementar la cantidad de proteína que se producía, lo cual fue una gran ventaja adicional”.
El objetivo inicial, sin embargo, no era desarrollar una vacuna, ha explicado Karikó. “Mi objetivo era utilizar el ARNm para codificar una proteína terapéutica que se pudiera administrar a un paciente con un ictus o un infarto de miocardio porque yo trabajaba en el campo de la cardiología y la neurocirugía, y quería evitar que se produjera una inflamación que pudiera empeorar la situación del enfermo”.
Para la galardonada, las vacunas contra la COVID-19 son sólo el principio de una revolución biomédica en ciernes. “Ahora que ya se ha demostrado la utilidad de esta técnica para desarrollar vacunas, estoy convencida de que pronto tendremos más para otras enfermedades. Además es una terapia especialmente barata porque la medicina se produce en tu propio cuerpo, tú mismo te conviertes en la fábrica de medicamentos. Las aplicaciones son infinitas”.
Karikó ha destacado que ya hay ensayos clínicos en fase avanzada de esta técnica contra enfermedades cardiovasculares, “introduciendo el ARNm en el corazón durante una cirugía de by-pass para mejorar la capacidad cardiaca”, explicó. “Pensábamos que esta sería la primera aplicación autorizada, pero la pandemia aceleró el desarrollo de las vacunas”.
También hay en marcha ensayos para probar vacunas de ARNm contra el VIH, la malaria y otras enfermedades, incluido el cáncer, según señala Karikó: “Hemos realizado ensayos prometedores con dos modelos animales para frenar una enfermedad autoinmune, la esclerosis múltiple. Se han iniciado muchos ensayos, y cada vez más compañías están probando esta tecnología”.
Weissman, por su parte, destaca también su trabajo actual en el posible desarrollo de “terapias génicas para la anemia falciforme, un trastorno con el que nacen 200.000 personas cada año. Esperamos poder tratarles con una sola inyección de ARNm que actuará sobre las células madres de la médula ósea, reparando su defecto genético y curando la enfermedad. Esto transformará la medicina”.
Un “ejemplo de perseverancia” ante el escepticismo inicial de la comunidad científica
Langer también se había enfrentado décadas atrás al mismo escepticismo que Karikó. Antes de que en 1974 lograra crear micro- y nanopartículas para encapsular grandes moléculas “la gente no creía que fuera posible”, ha recordado tras conocer el fallo del jurado. “Incluso después de publicado el resultado mucha gente me dijo que estaba mal, no lo creían. Los primeros nueve proyectos de investigación que solicité fueron rechazados, y no pude conseguir un trabajo en un departamento de ingeniería química, que es mi disciplina”. Langer se incorporó al MIT como profesor de Bioquímica Nutricional en 1978.
Su tecnología, sin embargo, ha resultado “absolutamente crítica” −afirma− para las terapias de ARNm. “Si el ARN se inyectara directamente, simplemente se destruiría. En cambio al ponerlo en estas pequeñas partículas lo proteges cuando lo inyectas en el cuerpo, y así sobrevive para poder trabajar”. Además, las partículas pueden modular la velocidad a la que se administra el ARN y en algunos casos también el lugar del cuerpo al que es transferido. “Esto permite un suministro muy preciso”, explica.
Langer es hoy uno de los científicos más citados a escala mundial, autor de más de un millar de patentes. Es co-fundador de la compañía Moderna, creadora de una de las vacunas de ARNm. Al demostrar el concepto de la encapsulación en partículas para transferir macromoléculas al cuerpo, Langer abrió un amplísimo abanico de posibilidades, exploradas por numerosos grupos de investigación. “Mucha gente se basó en nuestro trabajo de 1974, muchas empresas han estado desarrollando ideas; he ayudado a arrancar a algunas de ellas. Hoy en día las partículas se utilizan para tratar distintos tipos de cáncer, enfermedades mentales como la esquizofrenia y la adicción a los opiáceos. También en el tratamiento de la diabetes de tipo 2, en la prevención de hemorragias y diferentes enfermedades oculares, para aliviar el dolor y enfermedades cardiovasculares”.
Aún en plena pandemia, y con el enorme impacto de las vacunas contra la Covid-19, Langer considera “un honor increíble ganar este premio, tanto por las increíbles personas que lo han recibido en el pasado como por aquellas con las que voy a compartirlo”.
“Tanto Langer como Karikó y Weissman son un ejemplo de perseverancia”, afirma Oscar Marín. “Sufrieron múltiples rechazos por lo arriesgado de su investigación y la tendencia al cortoplacismo de la política científica; su triunfo nos hace pensar en lo difícil que es vaticinar qué va a funcionar en biología, y cuántos avances han podido quedar en el camino por no asumir riesgos”.
Jurado y Comité Técnico de Biología y Biomedicina
El jurado de esta categoría ha estado presidido por Angelika Schnieke, catedrática de Biotecnología de la Universidad Técnica de Múnich (Alemania), y ha contado como secretario con Óscar Marín, catedrático de Neurociencias y director del Centro de Trastornos del Neurodesarrollo en King’s College London (Reino Unido). Los vocales han sido Dario Alessi, director de la Unidad de Fosforilación y Ubiquitinación de Proteínas-MRC en la Universidad de Dundee (Reino Unido); Lélia Delamarre, científica principal en el Departamento de Inmunología del Cáncer de Genentech (Estados Unidos); Robin Lovell-Badge, jefe sénior de grupo y director del Laboratorio de Biología de las Células Madre y Genética del Desarrollo del Instituto Francis Crick (Reino Unido); Ursula Ravens, catedrática sénior del Instituto de Medicina Cardiovascular Experimental de la Universidad de Friburgo (Alemania); Ali Shilatifard, titular de la Cátedra Robert Francis Furchgott de Bioquímica y Pediatría de la Universidad Northwestern (Estados Unidos); y Bruce Whitelaw, catedrático de Biotecnología Animal y director interino del Instituto Roslin en la Universidad de Edimburgo (Reino Unido).
En cuanto al Comité Técnico de Apoyo del Consejo Superior de Investigaciones Científicas (CSIC), ha estado coordinado por M.ª Victoria Moreno, vicepresidenta adjunta de Áreas Científico-Técnicas del Consejo Superior de Investigaciones Científicas (CSIC), e integrado por José Félix de Celis Ibeas, profesor de investigación en el Centro de Biología Molecular Severo Ochoa (CBM); Dolores González Pacanowska, coordinadora del Área Global Vida y profesora de investigación en el Instituto de Parasitología y Biomedicina López Neyra (IPBLN); José Luis Martínez Menéndez, profesor de investigación en el Centro Nacional de Biotecnología (CNB); M. Isabel Medina Méndez, coordinadora adjunta del Área Global Vida y profesora de investigación en el Instituto de Investigaciones Marinas (IIM); e Isabel Varela Nieto, profesora de investigación en el Instituto de Investigaciones Biomédicas Alberto Sols (IIBM).
Entrevista de la Sra. Karico
Siempre quise desarrollar RNA mensajero, en primer lugar entender que un nucleótido modificado no es inflamatorio y cómo incorporar y hacer que el mRNA no fuera inmunogéni co inflamatorio al incorporar nucleósidos modificados específicamente pseudouridina al mensajero RNA este era el mensajero no inflamatorio y no inmunogénico.
Antes del desarrollo de la vacuna mensajero RNA se había ensayado para tratamiento de diferentes enfermedades para el corazón y enfermedades hepáticas por lo que ya están enprogreso y incluso antes del desarrollo de la vacuna,
El desarrolló de la terapia de ARN mensajerop or décadas y solo la ultima fase paso muy rápidamente pero todos los componentes estaban listos para la vacuna. Teniendo en cuenta que el mensajero de los nucleos RNA no puede incorporarse al genoma por lo que no hay precedentes de que eso podría pasar el mRNA es una plataforma que puede ser utilizado para desarrollar muchos tipos de medicamentos no solo las vacunas, sino mRNA que codifica proteínas terapéuticas y usarlos para tratar corazón, hígado y muchas otras enfermedades genéticas y ahora mismo ya también desarrollo parat erapia de genes donde la modificación del genoma puede ser introducida.
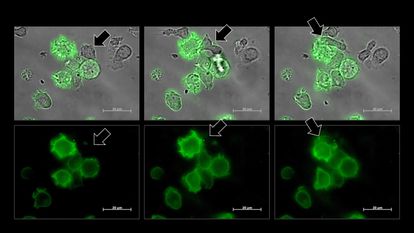 Una célula NK, señalada por una flecha, se vuelve verde tras robar un fragmento de la membrana de una célula cancerosa, también verde.HASIM ET AL.
Una célula NK, señalada por una flecha, se vuelve verde tras robar un fragmento de la membrana de una célula cancerosa, también verde.HASIM ET AL.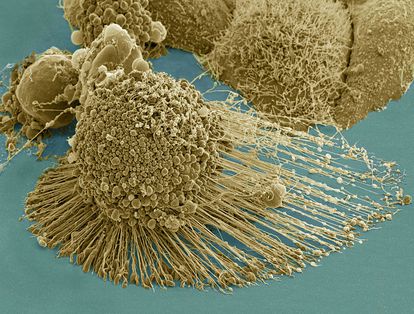

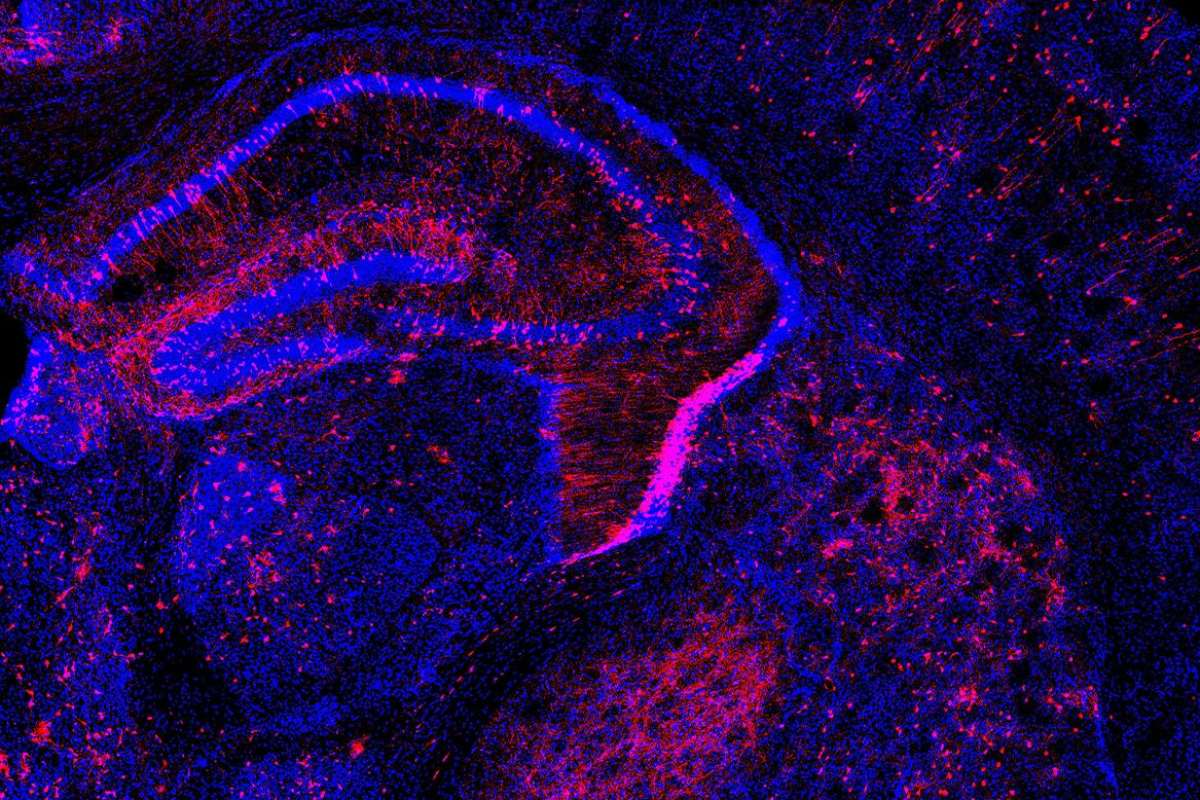 El método ‘Pertub-seq’ permite introducir cambios, como en esta célula de cerebro murino. Foto: LABORATORIO P. ARLOTTA (HARVARD)
El método ‘Pertub-seq’ permite introducir cambios, como en esta célula de cerebro murino. Foto: LABORATORIO P. ARLOTTA (HARVARD)






 pel
pel