El OMOMYC FREN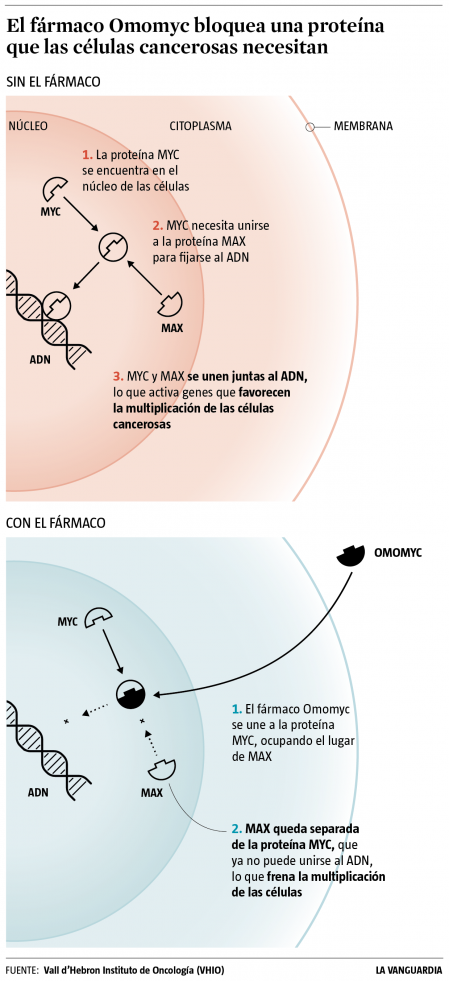 A AL CANCER
A AL CANCER
Posiblemente el cáncer ha existido siempre aunque sin duda alguna en un número muy inferior a la actualidad.
¿Es un problema de biología o de ambiente se puede concluir que es de los dos no cabe duda que el ambiente profesional que vivo genera cancer y también qué animales muy primitivos que vivían en ambiente poco polucionados también tenían cáncer pero en mucho menor número.
Lo cierto es que a medida que se tienen más medios para investigar se conocen mejor los mecanismos íntimos de esta enfermedad
Hace 76 millones de años, un dinosaurio herbívoro de la especie Centrosaurus apertus —. Murió junto a otros miembros de su manada, probablemente ahogado por una fuerte riada. Pero la enfermedad que padecía le habría terminado matando: osteosarcoma, cáncer de huesos, en estado avanzado.
También en 2020 se ha descrito un posible tipo raro de cáncer llamado histiocitosis de células de Langerhans en un hadrosaurio, un dinosaurio con pico que vivió en el Cretácico. Según los autores del estudio del centrosaurio, “las pruebas sugieren que los cánceres malignos, incluidos los de hueso, están profundamente arraigados en la historia evolutiva de los organismos”.
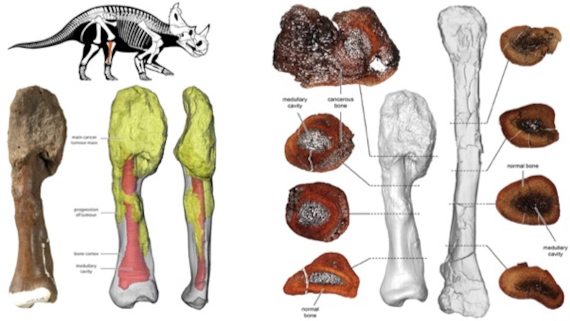 El peroné de Centrosaurus apertus, mostrando osteosarcoma. Crédito: Royal Ontario Museum/McMaster University
El peroné de Centrosaurus apertus, mostrando osteosarcoma. Crédito: Royal Ontario Museum/McMaster University
A pesar de ello, a menudo escuchamos un mensaje algo diferente: que el cáncer es una enfermedad humana moderna, causada por los factores ambientales nocivos a los que nos expone nuestro modo de vida.
En 2010, dos investigadores de las universidades de Manchester y Villanova esgrimían la escasa detección de cánceres en las momias egipcias para concluir que “el cáncer era raro en la antigüedad, y por tanto plantea preguntas sobre el papel de los factores ambientales carcinogénicos en las sociedades modernas”.
La egiptóloga Rosalie David, “no hay nada en el entorno natural que pueda causar cáncer. Así que tiene que ser una enfermedad hecha por el hombre, debida a la contaminación y los cambios en nuestra dieta y estilo de vida”.
El estudio provocó una oleada de comentarios de respuesta. Expertos en cáncer rebatían las palabras de David aportando ejemplos de factores naturales carcinogénicos, desde la luz ultravioleta del sol a compuestos químicos hallados en mohos y plantas.
En su blog de la revista Science, el químico Derek Lowe advertía contra lo que llamaba el efecto del Jardín del Edén: la idea de que “había un tiempo —hace mucho— en que la gente estaba en armonía con la naturaleza, comía alimentos naturales puros y saludables (del tipo de los que se supone que debemos comer), y no tenía todos los horribles problemas que tenemos en estos degenerados tiempos modernos”.
Lo cierto es que no solo las primeras descripciones del cáncer, e incluso su mismo nombre, se remontan a la antigüedad de la Grecia clásica, sino que se han encontrado signos de la enfermedad en fósiles de homininos africanos de hasta 1,7 millones de años de edad. Y en cuanto a los animales, la idea de que ciertas especies jamás padecen cáncer tiene más de mito que de realidad: incluso en el Ratopín o rata topo desnuda, el animal que suele esgrimirse como ejemplo de este fenómeno, se han encontrado casos. En estos peculiares animales, un estudio reciente sugiere que sus células no son intrínsecamente más resistentes al cáncer que las de otras especies, sino que posiblemente existan otros factores inmunitarios o del microentorno celular que las protegen. Es decir, hay una vulnerabilidad natural de las células animales al cáncer, que los propios mecanismos internos luchan por prevenir.
Por ejemplo, en los elefantes, una especie con una baja incidencia de cáncer, se ha encontrado que su genoma posee múltiples copias de p53, un gen supresor de tumores conocido como el “guardián del genoma”, por mantener a raya las mutaciones dañinas. Estos y otros mecanismos celulares son los que nos muestran que el cáncer, más que una consecuencia de nuestra sociedad, lo es de nuestra biología; por ejemplo, el osteosarcoma hallado en el centrosaurio suele darse en los humanos durante la adolescencia, cuando los huesos crecen rápidamente. Los supresores de tumores se encargan de vigilar, pero no son infalibles.
 En los elefantes, una especie con una baja incidencia de cáncer, se ha encontrado un gen supresor de tumores. Crédito: Diego Delso
En los elefantes, una especie con una baja incidencia de cáncer, se ha encontrado un gen supresor de tumores. Crédito: Diego Delso
“El cáncer no es una enfermedad moderna, sino que ha estado presente tanto en humanos como en animales de forma muy temprana desde el comienzo de la vida”, sentencia a OpenMind el paleopatólogo del hospital académico Bogenhausen de Múnich Andreas Nerlich. Este experto, que ha estudiado la incidencia del cáncer en poblaciones antiguas de Egipto y la Edad Media sin encontrar grandes diferencias con las de la era industrial, reconoce que hoy la enfermedad es mucho más frecuente, pero le atribuye una causa: “Mayor esperanza de vida, y por ello posiblemente una incubación más larga de sustancias carcinogénicas e infecciones virales con posible influencia carcinogénica”.
“El cáncer es una enfermedad propia de edades avanzadas”, remacha a OpenMind el investigador del Institute for Research in Biomedicine de Barcelona Manuel Serrano, experto en genes supresores de tumores. Serrano explica que esta es la causa de que en la antigüedad estas dolencias fueran menos frecuentes: “¡Llegar a los 50 también era muy raro! Antes llegaban pocos a los 80”. A ello se suma, añade Serrano, que antiguamente muchos casos pasaban inadvertidos: “Solo se diagnosticaba como cáncer aquel que se manifestaba externamente porque literalmente salía un bulto”.
Serrano apunta también que posiblemente los tipos de cánceres más frecuentes hayan cambiado respecto a tiempos más antiguos, dado que, abundando en el efecto del Jardín del Edén de Lowe, “la vida en la antigüedad no era la idealización que algunos tienen en mente”. “Se sabe que la gente inhalaba tóxicos en cantidades inimaginables debido a los fuegos que se hacían dentro de cuevas, chozas y cabañas”, expone. “Los niños, adultos y viejos, todos los días respiraban dentro de sus viviendas el humo del fuego”. Otro ejemplo que subraya Serrano son los alimentos, que en otros tiempos a menudo se consumían en mal estado. “Los alimentos podridos están llenos de bacterias y hongos que se sabe contienen agentes carcinogénicos”.
 Respuesta inmunitaria antitumoral. Crédito: NIH
Respuesta inmunitaria antitumoral. Crédito: NIH
Por último, destaca el investigador, los fármacos actuales nos liberan de infecciones crónicas que también suponen un factor de riesgo de cáncer.
OMOMYC CONTRA EL CÁNCER CREADO EN VALL D’HEBRON
Actúa de manera diferente a cualquier otra terapia y podría ser útil para múltiples tumores
Getty Images/Science Photo Libra Imagen creada por ordenador de células tumorales
El fármaco experimental Omomyc, que actúa contra el cáncer de una manera distinta a cualquier medicamento anterior y que tiene el potencial de mejorar el tratamiento de múltiples tumores, ha obtenido resultados positivos de eficacia y de seguridad en su primer ensayo clínico en pacientes.
El equipo del Instituto de Oncología de Vall d’Hebron (VHIO) que ha desarrollado el Omomyc está en conversaciones con empresas farmacéuticas para iniciar ensayos clínicos más amplios a mediados de 2023. Los primeros tipos de tumores en los que tiene previsto realizar estos ensayos son de páncreas, pulmón y mama triple negativo.
El Omomyc actúa inhibiendo la proteína MYC, que estimula la multiplicación de las células y que está hiperactivada en la mayoría de los cánceres. Se trata del primer fármaco oncológico dirigido contra una proteína del núcleo de las células, a diferencia de las decenas de terapias moleculares dirigidas contra proteínas de la membrana, a las que es más fácil acceder.
Los resultados obtenidos hasta ahora “nos dan motivos para ser optimistas”, valora Laura Soucek, investigadora del VHIO que empezó a trabajar en la proteína MYC hace 25 años y que ha persistido hasta que ha conseguido desactivarla. Entre investigadores del cáncer, MYC era calificada de undruggable -imposible de tratar con fármacos-.

Laura Soucek (izquierda) y Elena Garralda, en el Instituto de Oncología de Vall d’Hebron (VHIO)
Miquel González / Shooting / Colaboradores
En el ensayo clínico, iniciado en abril de 2021, han participado 22 pacientes con nueve tipos diferentes de cáncer. Todos ellos tenían metástasis, se les habían agotado las opciones de tratamiento y sus cánceres continuaban progresando. Se les administró Omomyc por vía endovenosa una vez por semana. El ensayo clínico se ha realizado en los hospitales Vall d’Hebron de Barcelona, Fundación Jiménez Díaz de Madrid y en el centro oncológico Clara Campal, también en Madrid.
Así funciona el Omomyc, el nuevo fármaco contra el cáncer
Como es habitual en los ensayos clínicos de fase 1, en que un nuevo tratamiento se administra por primera vez a personas, se empezó con dosis bajas y se fueron aumentando las cantidades hasta encontrar el punto de máxima eficacia con mínimos efectos secundarios.
Eficaz en casos distintos
La mejor respuesta se ha observado en un paciente con un tumor de glándulas salivales en los que la enfermedad está estabilizada desde hace 15 meses y que continúa en tratamiento, informa Elena Garralda, directora de la Unidad de Investigación de Terapia Molecular del Cáncer-CaixaResearch de Vall d’Hebron (UITM),que ha coordinado el ensayo clínico.
En otro paciente con sarcoma, se frenó la progresión del cáncer durante ocho meses. En pacientes con cáncer de páncreas, se estabilizó la enfermedad durante periodos de hasta seis meses, lo que ha supuesto un aumento de supervivencia para personas para las que no quedaba ningún otro tratamiento.
Cómo funciona el fármaco Omomyc
Según los resultados que se presentarán en un congreso sobre ensayos clínicos de cáncer que se inaugura hoy en Barcelona, el Omomyc ha conseguido que el cáncer dejara de progresar en aproximadamente la mitad de los pacientes. Concretamente, la enfermedad se estabilizó en ocho de los veintidós pacientes y no lo hizo en nueve, mientras que los otros cinco casos se consideran no evaluables (por ejemplo, porque murieron a los pocos días de iniciar el tratamiento).
El tratamiento ha sido bien tolerado por todos los pacientes excepto dos en los que ha sido necesario reducir la dosis debido a efectos secundarios.
En estos ensayos el Omomyc se administrará en combinación con otros tratamientos oncológicos. Si la terapia se inicia cuando la enfermedad no está tan avanzada, en un momento en que los pacientes aún tienen otras opciones de tratamiento, “esperamos que los resultados sean mejores que los del primer ensayo”, valora Soucek. Además, dado que el Omomyc inhibe la proliferación celular que favorece que el cáncer se vuelva resistente a los tratamientos, “esperamos conseguir un efecto de sinergia y que el Omomyc mejore la acción de otros fármacos”.
No se ha decidido aún con qué tipos de cáncer empezarán los ensayos clínicos de fase 2. Entre los primeros candidatos destaca el cáncer de páncreas, pues es un tipo de tumor en que la proteína MYC casi siempre está alterada, para el que faltan tratamientos eficaces y en el que los datos de los ocho pacientes tratados en el fase 1 han sido prometedores, informa Soucek. En este caso, el Omomyc se administraría en combinación con el tratamiento estándar de quimioterapia.
Un segundo candidato es el cáncer de pulmón no microcítico, que representa más del 80% de los cánceres de pulmón y que fue el primero en que se ensayó el Omomyc en ratones, con prometedores resultados que se presentaron en Science Translational Medicine. El nuevo fármaco se administraría entonces en combinación con terapias moleculares.
Finalmente, el equipo del VHIO espera poder realizar un ensayo clínico de fase 2 con cáncer de mama triple negativo, que es el tipo de cáncer de mama que dispone de menos opciones de tratamiento y para el que el Omomyc también ha obtenido resultados prometedores en ratones. Representa entre un 10% y un 15% de todos los cánceres de mama.
“Esperamos que toda esta investigación sirva para que nuestro fármaco llegue finalmente a los pacientes”, declara Soucek, que desarrolló el Omomyc con la colaboración de Marie-Eve Baulieu para que pudiera acceder al núcleo de las células. En una conversación con La Vanguardia en 2019, tras presentar sus primeros resultados positivos en ratones, la investigadora recordó que “todo el mundo decía que inhibir MYC era imposible. Allí donde otros veían un problema imposible, yo veía dificultades técnicas que esperaba que se pudieran resolver”.
Referencias
SOUCEK, MARIE-EVE BAULIEU desarrollaron omomyc para que pudiera acceder al núcleo de las células.
LAURA SOUCEK Y ELENA GARRALDA, en el instituto de oncología de vall d’hebron (vhio)
MIQUEL GONZÁLEZ / shooting / colaboradores
JOSEP CORBELLA Barcelona 26/10/2022 06:44actualizado a 26/10/2022 10:36
BOGENHAUSEN DE Múnich andreas nerlich openmind el paleopatólogo del hospital académico



 Tony
Tony
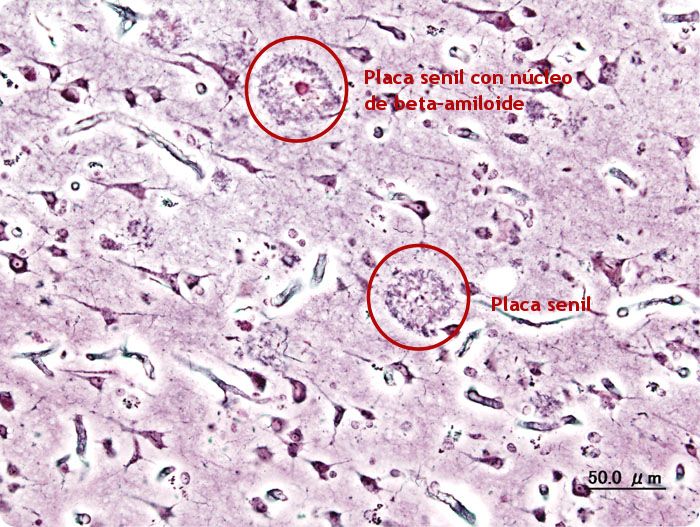
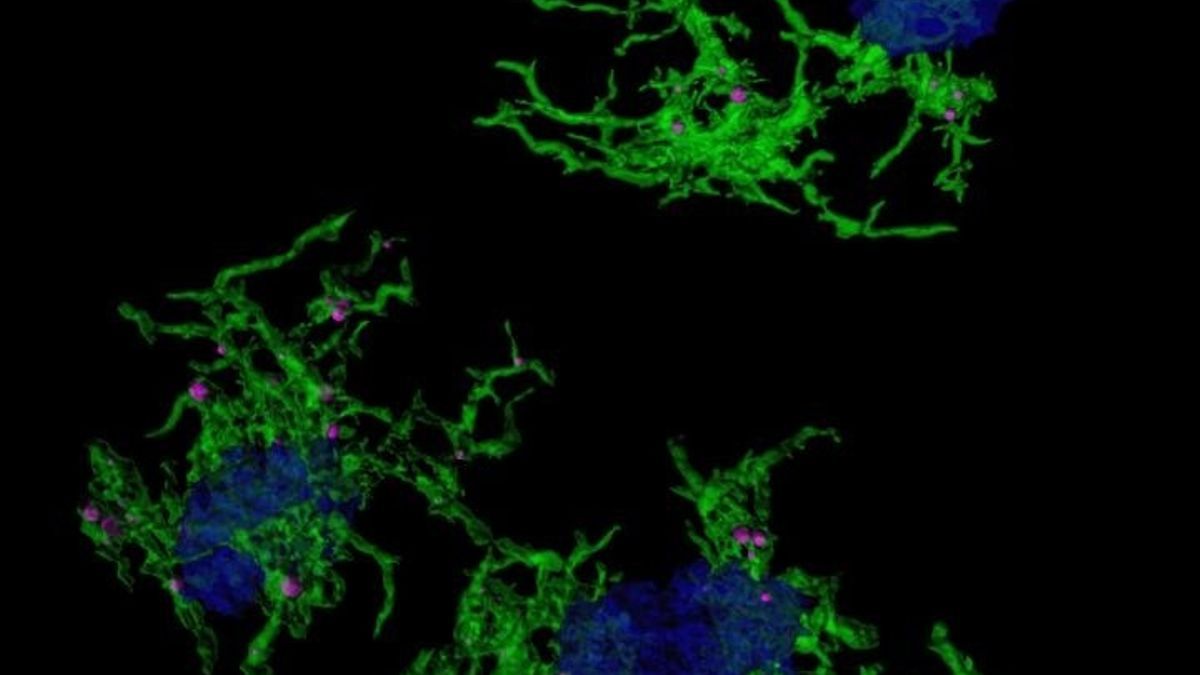 Imagen microscópica confocal microglia verde envuelve placas amiloide azul Europa Press
Imagen microscópica confocal microglia verde envuelve placas amiloide azul Europa Press

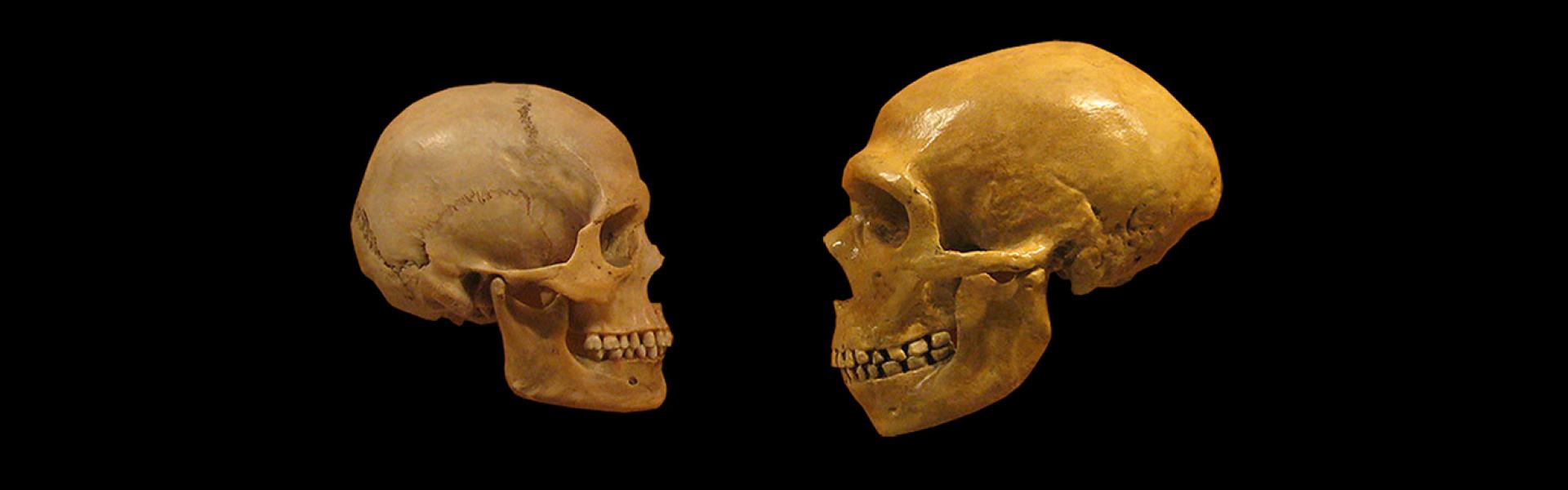
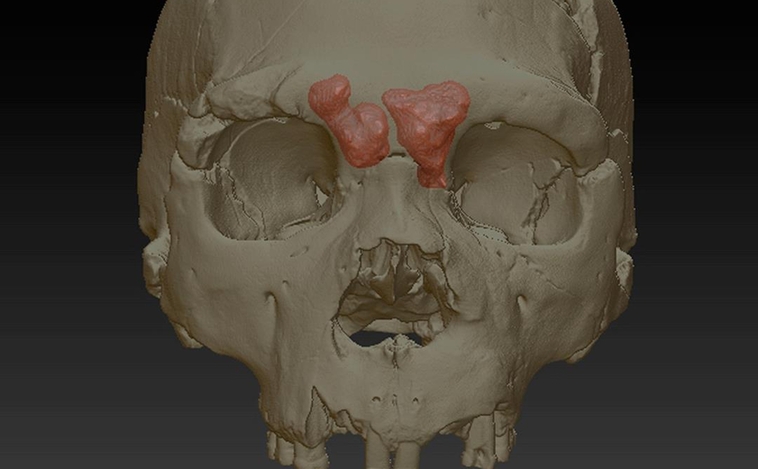





 Michael Eisenstein
Michael Eisenstein
 La condición de los seres vivientes más importante es la de lucha o huida y su persistencia conducen a la ansiedad en mayor o menor grado y es posiblemente la condición psíquica más frecuente y al mismo tiempo más difícil de combatir
La condición de los seres vivientes más importante es la de lucha o huida y su persistencia conducen a la ansiedad en mayor o menor grado y es posiblemente la condición psíquica más frecuente y al mismo tiempo más difícil de combatir