CELULAS DE LA ANSIEDAD EN UN CIRCUITO HIPOCAMPAL– HIPOTALAMICO
Este amplio estudio, enormemente complejo, se copia entero, y se deduce por su complejidad que son inimaginables, los mecanismos que utiliza el cerebro, para mantener los mecanismos de recompensa.
Es una revisión que se hace por:J C Jimenez, Katy su, A R Goldberg, L Paninski, R Hen, MQA Kheirbek
Los estímulos anxiogénicos se representan diferencialmente a lo largo del eje DV del HPC
La inhibición de la vHPC en ambientes ansiogénicos reduce el comportamiento de evitación
vCA1 se envía a LHA pero no a BA controla el comportamiento relacionado con la ansiedad
La mayoría de las neuronas de proyección vCA1-LHA representan estímulos ansiogénicos
Resumen
El hipocampo se piensa tradicionalmente para transmitir información contextual a las estructuras límbicas donde adquiere valencia. Usando imágenes de calcio y optogenética que se mueven libremente, mostramos que mientras la subregión CA1 dorsal del hipocampo se enriquece en el lugar de las células, el CA1 ventral (vCA1) se enriquece en las células de ansiedad que se activan en ambientes ansiogénicos y se requieren para el comportamiento de evitación. Las células de imagen definidas por su objetivo de proyección revelaron que las células de ansiedad estaban enriquecidas en la población vCA1 que se proyectaba al área hipotalámica lateral (LHA) pero no a la amígdala basal (BA). De acuerdo con esta selectividad, la activación optogenética de los terminales vCA1 en LHA pero no BA aumentó la ansiedad y la evitación, mientras que la activación de los terminales en BA pero no LHA afectó la memoria de miedo contextual. Así,
Introducción
El miedo y la ansiedad son respuestas emocionales a amenazas percibidas, con amenazas proximales que provocan miedo y amenazas distales que provocan ansiedad. En condiciones normales, los estados de ansiedad promueven comportamientos de evitación adaptativa que son críticos para navegar de forma segura en un entorno. La ejecución de comportamientos de evitación apropiados requiere el reconocimiento rápido de estímulos amenazadores y el enrutamiento de esa información a estructuras que puedan modular directamente estos comportamientos defensivos.
Si bien la evitación es adaptativa en condiciones normales, puede convertirse en una mala adaptación cuando las respuestas son excesivas e inapropiadas. En los seres humanos, una característica compartida de una serie de trastornos de ansiedad es la sobreestimación de la amenaza, que conduce a una mayor evitación (
Jovanovic y Ressler, 2010 Kheirbek et al., 2012 ). Sin embargo, los mecanismos y los circuitos neuronales por los cuales surgen conductas normales de evitación adaptativa y cómo estos circuitos se desordenan en enfermedades psiquiátricas, siguen siendo difíciles de alcanzar.
Si bien se sabe que el hipocampo (HPC) es crítico para los procesos cognitivos como la memoria episódica y la navegación espacial, también está implicado en la patogénesis del estado de ánimo y los trastornos de ansiedad. Una forma en que el HPC puede contribuir a los procesos cognitivos y relacionados con el estado de ánimo es a través de la heterogeneidad funcional a lo largo de su eje dorsoventral, con el HPC dorsal contribuyendo a funciones cognitivas como el aprendizaje y la memoria y el HPC ventral (vHPC) modulando la regulación emocional ( Fanselow y Dong, 2010 , Strange et al., 2014 ). Las lesiones del HPC ventral pero no dorsal son ansiolíticas, con un efecto mínimo en el aprendizaje espacial ( Bannerman et al., 2002 , Kjelstrup et al., 2002 , Moser et al., 1995), mientras que las lesiones dorsales de HPC afectan el aprendizaje espacial sin afectar las medidas relacionadas con la ansiedad. Además, las celdas de lugar, que se cree que contribuyen a una representación espacial del entorno, son más abundantes, estables y están sintonizadas en HPC dorsal en relación con la vHPC ( Ciocchi et al., 2015 , Jung et al., 1994 Keinath et al., 2014 , Royer et al., 2010 ). Además, estudios recientes de optogenética y farmacológicos indican que la manipulación de la vHPC o sus entradas y salidas corticales pueden afectar directamente el comportamiento relacionado con la ansiedad (Felix-Ortiz et al., 2013 Kheirbek et al., 2013 Kjaerby et al., 2016 Padilla-Coreano et al., 2016 Parfitt et al., 2017 Samuels et al., 2015. Wu y Hen, 2014 ).
A pesar de la evidencia acumulada que respalda el papel de la vHPC en los comportamientos anímicos y de ansiedad, poco se sabe acerca de cómo la vHPC representa información sobresaliente emocionalmente y cómo esas representaciones contribuyen al comportamiento. En el modelo de roedores, el comportamiento relacionado con la ansiedad puede evaluarse con tareas de evitación basadas en conflictos, que promueven el comportamiento de evitación adaptativa normal a amenazas distantes (Calhoon y Tye, 2015).
Por lo tanto, dilucidar cómo se representan los contextos ansiogénicos de forma innata en la vHPC será fundamental para comprender cómo puede guiar las conductas de evitación durante las tareas de ansiedad basadas en conflictos.
Ventral CA1 (vCA1) envía densas proyecciones a varias estructuras subcorticales como la amígdala basal (BA), el hipotálamo, el núcleo accumbens (NAc) y el núcleo del lecho de la estría terminal (BNST) (Canteras, 2002 Cenquizca y Swanson, 2006. Cenquizca y Swanson, 2007. Kishi et al., 2006 Tannenholz et al., 2014). Sin embargo, poco se sabe acerca de cómo vCA1 interactúa con estas regiones para organizar comportamientos emocionales. En particular, a pesar de décadas de trabajo que demuestran las interacciones hipocampal-hipotalámica en el amortiguamiento de las respuestas al estrés mediante la inhibición indirecta del eje hipotalámico-hipofisario-suprarrenal ( Jacobson y Sapolsky, 1991 Ulrich-Lai y Herman, 2009 ), la función de la vía hipotalámica HPC directa en el comportamiento modulador sigue endo desconocida. vCA1 envía directamente proyecciones densas a la BA y al área hipotalámica lateral (LHA), y estudios optogenéticos recientes han indicado que tanto la BA como la LHA pueden controlar el comportamiento relacionado con la ansiedad en tiempo real (Jennings et al., 2013 Tye et al., 2011). Sin embargo, aún no está claro cómo se representa la información relacionada con la ansiedad dentro de las neuronas de proyección vCA1 para impactar estas estructuras de salida.
Aquí, aplicamos imágenes de calcio y optogenética con libertad de movimiento para investigar cómo se representa la información relacionada con la ansiedad dentro de distintas poblaciones de neuronas de proyección vCA1.
De manera intrigante, encontramos «células de ansiedad» enriquecidas dentro de una población de neuronas proyectoras vCA1-LHA que representan ambientes ansiogénicos y causan un impacto causal en el comportamiento. Esto revela que el flujo de proyección vCA1-LHA puede servir como una ruta directa para que vCA1 controle rápidamente los comportamientos similares a la ansiedad.
Resultados
Representaciones de información relacionada con la ansiedad en vCA1
Primero determinamos cómo se activa vCA1 durante la exploración de entornos ansiogénicos de manera innata. Utilizamos la microendoscopia para realizar imágenes de calcio de las neuronas vCA1 que expresan GCaMP6f en ratones que se mueven libremente. Se implantó una lente de índice de refracción en gradiente (GRIN) sobre la subregión vCA1 ( Figuras 1 A y S1 A), y el indicador de Ca 2+ GCaMP6f se expresó de forma viral para visualizar la actividad de Ca 2+ devCA1 como se describió anteriormente (Resendez et al., 2016. Ziv et al., 2013 ). Este enfoque nos permitió registrar eventos transitorios de Ca 2+ en neuronas vCA1 individuales en el mismo campo de visión (FOV), mientras que los ratones exploraron libremente múltiples entornos ( Figura 1 A; consulte Métodos STAR ).
Figura 1 Representaciones de información relacionada con la ansiedad en vCA1
En el laberinto más elevado (EPM), encontramos que la mayoría de las neuronas vCA1 mostraron un aumento significativo en la actividad de Ca 2+ y en la tasa de transitorios de Ca 2+ durante la exploración del compartimento ansiogénico de brazo abierto en comparación con el compartimiento de brazo cerrado ( Figuras 1 B, 1C, S1 B y S1D). A continuación, comparamos la actividad de calcio promedio a través de entradas de comportamiento sucesivas y episodios de exploración de los brazos abiertos. Encontramos que la actividad de vCA1 aumenta durante la exploración de ambos brazos abiertos y disminuye al reingresar al compartimiento de brazos cerrados ( Figura 1RE). Este aumento de la actividad no fue impulsado simplemente por un cambio en la ubicación espacial, ya que el cambio entre los compartimentos de brazo cerrado no provocó una mayor actividad, mientras que la actividad mayor sostenida de cambio de brazo abierto ( Figura 1D). Además, esto no se debió a las diferencias en la velocidad del ratón entre los compartimentos, ya que las distribuciones de velocidad del brazo abierto y del brazo cerrado fueron similares entre los animales ( Figura S1 C). A continuación, consideramos si la actividad aumentada de vCA1 con el brazo abierto estaba relacionada con un aumento de la importancia espacial en el brazo abierto (en relación con el brazo cerrado), en lugar de su naturaleza aversiva. Imaginamos vCA1 mientras los ratones exploraban un campo de campo abierto familiar que incluía un objeto novedoso espacialmente sobresaliente que provocaba un enfoque ( Figura 1MI). A diferencia de nuestros hallazgos en los brazos abiertos del EPM, la exploración del cuadrante que contiene el objeto novedoso apetitivo no evocó aumentos en la actividad de vCA1 ( Figura 1 F), lo que indica que las neuronas vCA1 están sesgadas para representar características ansiogénicas del entorno en lugar de cambios en saliencia espacial.
A continuación, investigamos si la magnitud de la actividad evocada con el brazo abierto estaba correlacionada con el nivel de ansiedad de los animales individuales. La actividad evocada de brazo abierto vCA1 ( tasa transitoria de Ca 2+ abierto-cerrado ) se correlacionó estrechamente con el grado en que los ratones evitaban los brazos abiertos del laberinto (una medida de los niveles de ansiedad de referencia)), con más Animales ansiosos que exhiben mayores niveles de actividad. Es importante destacar que estos efectos en las diferencias de frecuencia de Ca 2+ de brazo abierto y cerrado no fueron un artefacto de muestreo de comportamiento escaso, ya que el cálculo de las tasas de eventos de Ca 2+ de brazo cerrado a partir de un número de muestras de comportamiento coincidente dio lugar a la misma correlación entre Actividad evocada por el brazo y evitación de los brazos abiertos. Además, mientras que la tasa media de eventos de Ca 2+ en el brazo abierto se correlacionó positivamente con el comportamiento de evitación, las tasas del brazo cerrado no se correlacionaron con la evitación del brazo abierto y S1H). Finalmente, se descubrió que la actividad del brazo abierto vCA1 aumenta aún más cuando los ratones se involucran en comportamientos de inclinación de cabeza altamente ansiogénicos en los bordes de los brazos abiertos. Estos resultados sugieren que vCA1 genera representaciones de estímulos ansiogénicos a través de un código de frecuencia que se correlaciona con el estado de ansiedad de referencia y las escalas con la naturaleza aversiva de la conducta.
Control en tiempo real del comportamiento de evitación por vCA1
A continuación, probamos si la actividad evocada de brazo abierto vCA1 era necesaria para el mantenimiento de la evitación de brazo abierto en el EPM. Los ratones se inyectaron bilateralmente en vCA1 con un virus de control o uno que expresaba ArchT y se implantaron con fibra óptica en la misma ubicación. Esto nos permitió silenciar las neuronas piramidales que expresan vCA1-ArchT con iluminación de luz de 532 nm . Activamos selectivamente el silenciamiento optogenético de la actividad de vCA1 solo cuando los ratones ingresaron a los brazos abiertos del EPM. Cuando se compararon con los controles de eYFP, los ratones silenciados con vCA1-ArchT pasaron significativamente más tiempo explorando los brazos abiertos del EPM. Este efecto también se encontró en la prueba de campo abierto (OFT), ya que el silenciamiento selectivo de las neuronas vCA1 durante la exploración de la zona central ansiogénica aumentó significativamente la cantidad de tiempo que los ratones pasaron explorando el centro. Esto no se debió a los efectos apetitivos de la inhibición de vCA1 inducida por la luz, ya que los ratones Arch y eYFP pasaron una cantidad similar de tiempo en el lado con iluminación de luz en un ensayo de preferencia de lugar en tiempo real (RTPP). Además, estos efectos no se debieron a cambios en la actividad locomotora o aumentos en el número de visitas de brazo abierto. Para evaluar si estos cambios en el comportamiento relacionado con la ansiedad fueron específicos para silenciar la actividad evocada con el brazo abierto o debido a la incapacidad de los ratones para reconocer dónde estaban en el laberinto, luego probamos un segundo grupo de ratones con estimulación con láser solo durante el período cerrado. – Combates de exploración del brazo. En contraste con el silenciamiento de brazo abierto en el EPM, el silenciamiento de brazo cerrado no causó cambios en el comportamiento de ansiedad. Estos datos sugieren que la actividad aumentada de vCA1 en ambientes ansiogénicos promueve el comportamiento de evitación.
Representaciones diferenciales de información relacionada con la ansiedad en el eje dorsoventral de CA1
A continuación, evaluamos la especialización y la estabilidad de la actividad relacionada con la ansiedad a lo largo del eje dorso-ventral del hipocampo. Se tomaron imágenes de las neuronas dCA1 en el EPM de una manera idéntica a la descrita anteriormente y se comparó la actividad con las neuronas con imagen vCA1. A nivel poblacional, encontramos que las neuronas dCA1 no mostraron cambios significativos en la tasa de eventos transitorios de Ca 2+ en los brazos abiertos del EPM y la actividad de dCA1 no se correlacionó con el nivel de ansiedad de los animales individuales. Uso de métodos estadísticos para clasificar las neuronas como significativamente más activas en compartimientos selectivos, encontramos que ∼51% de las neuronas vCA1 registradas eran de brazo abierto, y esta proporción fue significativamente mayor en relación con la población de dCA1 que exhibió una preferencia de selectividad distribuida más uniformemente.
Figura 3 Representaciones diferenciales de información relacionada con la ansiedad a lo largo del eje dorsoventral de CA1
A continuación, consideramos si las neuronas selectivas de brazo abierto CA1 en la EPM estaban especializadas para responder a estímulos ansiogénicos en contextos múltiples. Las neuronas vCA1 y dCA1 individuales se rastrearon a través de múltiples sesiones de imágenes, y se evaluó la actividad de las neuronas de brazo abierto EPM en la OFT que provoca ansiedad y la tarea de objeto novedoso apetitivo. De manera interesante, encontramos que las neuronas selectivas de brazo abierto vCA1 exhibieron una tasa significativamente mayor de eventos transitorios de Ca 2+durante la exploración de la zona central ansiogénica de la OFT en comparación con la periferia, pero no durante la exploración de un objeto novedoso preferido .
Además, estos efectos fueron específicos de las neuronas que prefieren el brazo abierto vCA1, ya que las neuronas del brazo abierto dCA1 no mostraron cambios en la actividad de Ca 2+ en el centro OFT. En un enfoque imparcial alternativo, definimos las neuronas selectivas de tarea vCA1 y dCA1 en las tareas EPM, OFT y objeto nuevo según su preferencia de actividad para los compartimentos ansiogénico (brazos abiertos; centro) o apetitivo (zona objeto nuevo) y comparamos los superposición de celdas selectivas reclutadas en todas las tareas. Encontramos que la población de neuronas vCA1 que eran selectivas para los brazos abiertos EPM se superponían significativamente con neuronas que eran selectivas para el centro OFT, pero no con neuronas selectivas a un objeto nuevo, lo que indica que las neuronas selectivas vCA1-brazo abierto son Preferentemente reclutados en ambientes ansiogénicos. En contraste, las neuronas dCA1 que eran selectivas para los brazos abiertos EPM no se superponían con las neuronas dCA1 OFT centradas en el centro por encima de los niveles de probabilidad ( Figura S3H), pero fueron reclutados preferentemente para un objeto novedoso apetitivo. Esto indica que las neuronas dCA1 que son selectivas para los brazos abiertos no son selectivas para ambientes ansiogénicos, sino que pueden ser sensibles a la novedad. Estos resultados sugieren la existencia de células que muestran representaciones estables de información relacionada con la ansiedad o «células de ansiedad» que son más abundantes en vCA1 que en dCA1.
Como nuestra estrategia de direccionamiento viral no pudo distinguir entre las células piramidales vCA1 y las interneuronas inhibitorias, se tomaron imágenes de las interneuronas inhibitorias vCA1 para determinar si las células de ansiedad estaban representadas en exceso en esta población. Expresamos viralmente un GCaMP6f dependiente de Cre en ratones vGAT-Cre (Vong et al., 2011) e imágenes de respuestas neuronales en la EPM. Encontramos que, en general, las neuronas vCA1-vGAT no exhibieron una mayor actividad en el compartimiento de brazo abierto, sino que la mayoría de las neuronas con imagen vCA1-vGAT eran preferenciales de brazo cerrado ( Figura 4 C). Estos resultados sugieren que las neuronas que prefieren el brazo abierto vCA1 se componen en gran parte de neuronas piramidales glutamatérgicas, en lugar de interneuronas inhibitorias.
Luego evaluamos si las neuronas vCA1 y dCA1 también podrían representar diferencialmente información espacial. Los ratones se tomaron imágenes en el mismo FOV mientras exploraban dos contextos con señales espaciales diferentes (contextos A y B), seguidos de una segunda exposición al contexto A (ABA), y se generaron mapas de velocidad de sus campos de disparo dentro de los contextos como se describió anteriormente . Encontramos que las neuronas dCA1 codificaban más información espacial Skaggs et al., 1996 y tenía campos de posición más estables en relación con vCA1, lo que indica que las neuronas dCA1 están más sintonizadas espacialmente que vCA1. Tomados en conjunto, estos datos sugieren que dCA1 está enriquecido en células de lugar, mientras que vCA1 está enriquecido en células de ansiedad.
Las neuronas de proyección del hipotálamo lateral vCA1 no se superponen significativamente con las proyecciones vCA1 a la amígdala basal o la corteza prefrontal medial
Dada la heterogeneidad de las respuestas de vCA1 en el EPM, a continuación identificamos a través de qué flujos subcorticales de vCA1 se pueden mediar estos efectos sobre el comportamiento relacionado con la ansiedad. Nos enfocamos en dos estructuras que reciben algunas de las proyecciones vCA1 más densas y se sabe que contribuyen al comportamiento relacionado con la ansiedad, el miedo aprendido y las respuestas al estrés: la amígdala basal (BA) y el hipotálamo lateral (LHA) (Canteras y Swanson, 1992 , Cenquizca y Swanson, 2006. Kishi et al., 2006 , Tannenholz et al., 2014).
La inyección de virus CaMKII-ChR2-eYFP en vCA1 reveló un marcaje terminal denso en el BA (dentro de la amígdala basomedial y basolateral) y el LHA con intensidades similares. Las grabaciones de cortes agudos en neuronas BA o LHA en ratones inyectados con eYFP ChR2 confirmaron que la entrada monosináptica de vCA1 a BA y LHA, ya que la estimulación con láser a 473 nm dentro de los subcampos BA o LHA fue suficiente para provocar corrientes glutamatérgicas, monosinápticas, excitadoras y sinápticas (EPSC) .
A continuación, determinamos si diferentes poblaciones de neuronas vCA1 se proyectan a LHA y BA, como se vio recientemente con otras salidas a BLA, CeA, mPFC, septum lateral y NAc ( Cembrowski et al., 2016 , Jin y Maren, 2015 , Kim y Cho, 2017 , Lee et al., 2014b, Okuyama et al., 2016, Parfitt et al., 2017 Xu et al., 2016 ). La inyección de los trazadores retrógrados de la subunidad B de la toxina del cólera (CTB) CTB-555 y CTB-488 en el BA y LHA reveló que estos proyectores vCA1 eran en gran medida poblaciones no superpuestas, ya que solo el 3% de las neuronas etiquetadas enviaron proyecciones duales. De manera interesante, también encontramos que las neuronas proyectoras BA y LHA se segregaron anatómicamente y se organizaron de manera laminar, con neuronas vCA1-LHA ubicadas más profundamente en la capa piramidal CA1 en relación con las neuronas vCA1-BA. A continuación, evaluamos si las neuronas proyectoras de VCA1-LHA envían colaterales a la corteza prefrontal medial (mPFC), una vía que recientemente se ha descrito para modular el comportamiento relacionado con la ansiedad (Padilla-Coreano et al., 2016). Realizamos estudios de rastreo retrógrado similares a los descritos anteriormente e inyectamos CTB-555 en el mPFC y CTB-488 en el LHA y encontramos que, de forma similar a las vías vCA1-BA, las neuronas proyectantes vCA1-LHA no se superponían con las neuronas proyectoras vCA1-mPFC. En conjunto, estos estudios indican que las neuronas que proyectan vCA1-LHA surgen de poblaciones de células en gran medida no superpuestas en relación con las proyecciones de vCA1-BA y vCA1-mPFC y ocupan capas anatómicamente distintas dentro de vCA1.
Las proyecciones vCA1-Amygdala y vCA1-LHA contribuyen de manera diferencial al comportamiento relacionado con la ansiedad y las memorias contextuales de miedo
Dada la segregación anatómica de las proyecciones vCA1-BA y LHA, a continuación determinamos si la modulación de los proyectores vCA1-BA y vCA1-LHA contribuyen de manera diferencial al comportamiento. A los ratones se les inyectó ChR2-eYFP o un virus eYFP de control en vCA1, y se implantaron fibras ópticas en la amígdala (dirigida a los núcleos basales) o LHA. Luego, a los ratones se les hicieron pruebas de efectos de luz en pruebas de comportamiento relacionado con la ansiedad y condicionamiento del miedo contextual (CFC).
Los proyectores vCA1-Amygdala y vCA1-LHA contribuyen de manera diferencial al comportamiento relacionado con la ansiedad y al miedo aprendido
En CFC, los ratones exploraron el contexto de condicionamiento mientras recibían la estimulación con láser, después de lo cual recibieron un breve shock en el pie. El día dos, los ratones se colocaron de nuevo en el mismo contexto en ausencia de estimulación con láser para probar los efectos de luz en la codificación de CFC. Encontramos que los ratones vCA1-amígdala-ChR2 se congelaron significativamente menos que los controles, lo que indica que la interrupción de los patrones de actividad normal entre vCA1 y la amígdala fue suficiente para interrumpir la codificación del miedo contextual. Para determinar si se requirieron patrones de actividad vCA1-BA intactos para la recuperación, una cohorte diferente de ratones se entrenó con luz apagada y se probó la congelación en el día dos con la luz encendida. Los ratones vCA1-amígdala-ChR2 se congelaron menos que los controles, lo que indica que la actividad de vCA1-amígdala era necesaria tanto para la codificación como para la recuperación de la memoria de miedo contextual. Estos efectos se recapitularon con la inhibición del terminal vCA1-amígdala, lo que indica que los efectos de la excitación de ChR2 probablemente se debieron a una pérdida de la función. Sorprendentemente, realizar las mismas manipulaciones en la vía vCA1-LHA no afectó ni a la codificación ni a la recuperación del miedo contextual, y estos efectos negativos no fueron específicos de la frecuencia, que indica un rol selectivo para vCA1-amígdala en la codificación y recuperación del contexto.
A continuación, probamos la contribución de estas proyecciones de vCA1 al comportamiento relacionado con la ansiedad. Si bien el silenciamiento o la estimulación de los terminales vCA1-amígdala no tuvo ningún efecto sobre el porcentaje de la distancia al centro en la OFT), la estimulación disminuyó fuertemente la exploración del centro en los ratones vCA1-LHA-ChR2, un efecto que persiste La siguiente época de la luz apagada. Además, en un ensayo RTPP, mientras que la estimulación no produjo un efecto en los ratones vCA1-amígdala-ChR2, la estimulación en los ratones vCA1-LHA-ChR2 provocó evitación, como expresión de vCA1-LHA-ChR2 los ratones pasaron significativamente menos tiempo en la cámara de estimulación en relación con los controles. Es importante destacar que los efectos de la luz en los ratones vCA1-LHA-ChR2 no se debieron a cambios en la actividad locomotora. Estos estudios indican que la modulación de las neuronas de proyección vCA1-LHA pero no vCA1-amígdala puede afectar los comportamientos relacionados con la ansiedad y provocar evitación.
Estos resultados apoyan una disociación funcional entre las neuronas vCA1-amígdala y vCA1-LHA, con proyecciones de vCA1-BA que modulan la recuperación y codificación de la memoria de miedo contextual y las neuronas vCA1-LHA conducen el comportamiento y la aversión relacionados con la ansiedad.
La proyección vCA1-LHA está enriquecida en células de ansiedad
A continuación, investigamos si esta disociación funcional ya estaba presente en el nivel de vCA1. Se inyectó un virus adeno tipo 2-Cre canino retrógrado (CAV2-Cre) en el subcampo BA o LHA, y se inyectó un virus GCaMP6f dependiente de Cre en vCA1. Luego se implantó una lente GRIN sobre la región vCA1, y se realizó una imagen de la actividad de Ca 2+ específica de la proyección durante condiciones de comportamiento idénticas a las descritas anteriormente. Utilizando este enfoque, los terminales vCA1-GCaMP6f se visualizaron selectivamente en los subcampos BA o LHA, pero no en ambos, confirmando la expresión específica de proyección del indicador de Ca 2+ .
La actividad de imagen en el EPM en las dos poblaciones reveló que, si bien las neuronas vCA1-BA y vCA1-LHA exhibían una mayor actividad en los brazos abiertos, la magnitud de la diferencia entre la actividad del brazo abierto y la del brazo cerrado fue mayor en las neuronas vCA1-LHA en relación con las neuronas que proyectan vCA1-BA. Además, encontramos que las neuronas vCA1-LHA estaban altamente enriquecidas en células de ansiedad en relación con las neuronas vCA1-BA, con células de ansiedad que representan el 79% de la población de vCA1-LHA. Es importante destacar que los grupos no difirieron significativamente en el porcentaje de tiempo de brazo abierto y la actividad de Ca 2+ en el compartimiento de brazo cerrado EPM no difirió según el tipo de proyección.
A continuación, analizamos la estructura de ajuste espacial y la información espacial de las poblaciones proyectadas como se describió anteriormente y no encontramos diferencias significativas entre las poblaciones de proyección vCA1-BA y vCA1-LHA Estos estudios sugieren que si bien las neuronas que proyectan vCA1-LHA están enriquecidas en respuestas de actividad relacionadas con la ansiedad en relación con los proyectores vCA1-BA, ambas corrientes de proyección codifican bajos niveles de información espacial.
Teniendo en cuenta el enriquecimiento de las células de ansiedad dentro de la vía vCA1-LHA, a continuación evaluamos si la actividad en estas neuronas era necesaria para evitar el comportamiento como se probó previamente en nuestras manipulaciones de población total. Inyectamos CAV2-Cre en la LHA y un ArchT dependiente de Cre o virus de control en vCA1 e implantamos fibra óptica en vCA1 ( Figuras 7 F y S6 H). Probamos ratones en el EPM, y activamos selectivamente la estimulación con láser cuando los ratones ingresaron al compartimiento de brazo abierto para silenciar la actividad de las células de ansiedad vCA1-LHA. Encontramos que el silenciamiento de las neuronas vCA1-LHA durante la exploración de brazos abiertos redujo significativamente la evitación de los brazos abiertos, recapitulando los efectos que encontramos en la manipulación de toda nuestra población.
Estos resultados apoyan la hipótesis de que la información de valencia negativa se representa a nivel de vCA1, se enriquece en las neuronas que se proyectan hacia el LHA y es necesaria para el comportamiento de evitación.
Discusión
Una representación de contextos anxiogénicos en vCA1
El HPC integra información sensorial diversa de la corteza entorrinal (CE) para generar representaciones complejas del entorno (
Canto et al., 2008 ), que puede combinarse con estímulos aversivos para apoyar el condicionamiento del miedo contextual (
Kim y Fanselow, 1992 Phillips y LeDoux, 1992). En contraste con esta visión puramente cognitiva de la HPC, nuestros estudios de imagen de vCA1 indican que las neuronas de la vHPC tienen una representación de estímulos ansiogénicos innatos. Encontramos que vCA1 está enriquecido en células de ansiedad que responden al compartimiento de brazo abierto del EPM. Además, estas neuronas son reclutadas preferentemente por otros entornos ansiogénicos, como el centro de la OFT, pero no por un nuevo objeto apetitivo. Además, nuestros experimentos de silenciamiento optogenético de circuito cerrado demuestran que este aumento en la actividad de vCA1 en contextos ansiogénicos es necesario para la expresión del comportamiento de evitación. Curiosamente, encontramos que las células de ansiedad vCA1 aumentan en gran medida su actividad después de la entrada en los brazos abiertos ansiogénicos del EPM, Jacinto et al., 2016 ]). Por lo tanto, vCA1 puede modular los comportamientos de ansiedad mediante la codificación directa de estímulos amenazadores a través de células de ansiedad que están especializadas para representar ambientes ansiogénicos innatos.
Esta representación ansiogénica puede originarse dentro del circuito de HPC o puede proporcionarse mediante entradas extra-hipocampo. Uno de esos aportes es el BLA, que se proyecta directamente a vCA3 y vCA1, y cuyos aportes han demostrado recientemente que afectan los comportamientos relacionados con la ansiedad (Felix-Ortiz et al., 2013 ). Sin embargo, esto se complica con las grabaciones de una sola unidad, que han demostrado que las neuronas BA son preferentemente activas en el compartimiento seguro de los brazos cerrados del EPM, en contraste con las células de ansiedad vCA1 ( Adhikari et al., 2015 Wang et al., 2011). Además, se encontró que las neuronas BLA que se proyectan a vCA1 responden a señales de valencia positiva y negativa ( Beyeler et al., 2016 ), en lugar de desviarse hacia estímulos de valencia negativos como en las células de ansiedad vCA1.
Alternativamente, esta representación ansiogénica en vCA1 podría surgir a través de entradas de la CE especializadas para reconocer características ambientales específicas que contribuyen a contextos ansiogénicos como el cambio de color, las diferencias en la iluminación, la elevación y la falta de paredes ( Diehl et al., 2017
Lu et al., 2013 ). Por lo tanto, las representaciones de valencia innata podrían surgir a través del enrutamiento selectivo de estas características a las poblaciones de vCA1 definidas por proyección que modulan el comportamiento de evitación. Por lo tanto, los estudios futuros que registran y controlan las regiones de entrada determinarán la contribución relativa de los circuitos EC, BLA y CA3 ascendente y del giro dentado en la generación de la señal ansiogénica en vCA1.
Corrientes de proyección divergentes vCA1
Recientemente se ha apreciado que el vCA1 envía proyecciones paralelas y en gran parte no superpuestas a mPFC, septum lateral, NAc, BLA y amígdala central (Ce) ( Cembrowski et al., 2016 Jin y Maren, 2015 Kim y Cho, 2017 Lee et al., 2014b Okuyama et al., 2016 Parfitt et al., 2017 , Xu et al., 2016), y aquí encontramos esta segregación también dentro de las vías de BA y LHA. De manera interesante, en contraste con las proyecciones vCA1 entremezcladas a BA y Ce, encontramos que las neuronas de proyección vCA1-BA y vCA1-LHA se organizaron de manera laminar dentro de CA1. Estudios recientes sobre la laminación de CA1 han demostrado que las neuronas piramidales en las capas profundas y superficiales de CA1 difieren en sus propiedades fisiológicas, entradas inhibitorias locales e entradas de largo alcance (Danielson et al., 2016b , Lee et al., 2014b , Li et al., 2017, Masurkar et al., 2017). Por lo tanto, las neuronas proyectoras vCA1-BA y vCA1-LHA pueden recibir diferentes entradas y exhibir diferentes propiedades fisiológicas, lo que permite el enrutamiento de información diferencial entre estas poblaciones.
vCA1-LHA como una ruta directa al comportamiento de evitación de control
Nuestros estudios revelaron un sorprendente control específico de la vía del comportamiento similar a la ansiedad en vCA1, ya que la activación de los terminales vCA1-LHA pero no vCA1-BA genera un comportamiento de evitación en las tareas de ansiedad. Además, nuestros estudios de imágenes de proyección específica mostraron un enriquecimiento de las células de ansiedad dentro de las proyecciones de vCA1-LHA en relación con las proyecciones de vCA1-BA, con aproximadamente el 80% de la población de vCA1-LHA con una representación de los brazos abiertos del EPM. El LHA puede entonces integrar esta representación de vCA1 con las que surgen de otras áreas, como el BNST, que se encontró que tenía una representación de los brazos cerrados del EPM, y envía una proyección directa al LHA que produce efectos ansiolíticos (Kim et al., 2013 ). Por lo tanto, el enrutamiento de las representaciones de ansiedad al LHA puede ser crítico para la generación de conductas de evitación.
Aunque apuntamos al LHA en nuestros estudios, vCA1 envía proyecciones directas a varios subnúcleos del hipotálamo ( Canteras y Swanson, 1992 ), muchos de los cuales contienen diversos tipos de células y son anatómicamente difíciles de seleccionar selectivamente (Canteras, 2002 ). Estudios elegantes han comenzado a analizar estos diversos circuitos hipotalámicos ( Jennings et al., 2013 Jennings et al., 2015 Kunwar et al., 2015 , Lee et al., 2014a , Lin et al., 2011 , Silva et al., 2013 ), y los estudios futuros que investigan los tipos de células hipotalámicas a través de los cuales vCA1 y otros insumos provocan efectos en las conductas de evitación serán críticos para comprender cómo esta estructura modula las conductas similares a la ansiedad.
Nuestros estudios revelan proyecciones de vCA1 a la LHA como una nueva vía por la cual la vHPC puede modular el comportamiento relacionado con la ansiedad. Estudios recientes indican que las neuronas de proyección de vHPC-mPFC también representan información relacionada con la ansiedad en el EPM y que la inhibición optogenética de las entradas de vHPC al mPFC reduce la evitación de brazos abiertos (Ciocchi et al., 2015 ). Mientras que las poblaciones de células glutamatérgicas en el LHA pueden conducir la evitación rápida y los comportamientos aversivos directamente ( Hakvoort Schwerdtfeger y Menard, 2008 Jennings et al., 2013 Kim et al., 2013), el mPFC probablemente modula los comportamientos relacionados con la ansiedad a través de salidas a la amígdala, el hipotálamo, el tálamo o el gris periacueductal ( Do-Monte et al., 2015 , Likhtik et al., 2014 , Radley et al., 2006 , Sesack et al., 1989 , Sotres-Bayon y Quirk, 2010). Una posibilidad intrigante es que la contribución de las vías vCA1-LHA y vCA1-mPFC al comportamiento relacionado con la ansiedad es análoga al “camino bajo” talámico y al “camino alto” cortical en el control del condicionamiento del miedo auditivo dependiente de la amígdala (LeDoux, 1996, LeDoux, 2000 ). En este modelo, el enrutamiento diferencial de las representaciones contextuales ansiogénicas entre las vías de vCA1 podría cumplir una función similar de soportar tanto una señal de evitación rápida (mediante el enriquecimiento de señales ansiogénicas en proyecciones directas de vCA1-LHA) como una representación más lenta y de mayor orden mediante la integración cortical mediante La vía vCA1-mPFC.
Las neuronas de proyección vCA1-BA median el miedo aprendido, pero el comportamiento no evasivo innato
Un aspecto interesante de nuestros estudios fue que se observaron menos células de ansiedad en la población de vCA1-BA en comparación con la vía de vCA1-LHA, y la manipulación de las proyecciones de vCA1 a BA no tuvo impacto en las tareas de ansiedad innata, a pesar de que los estudios demuestran que la vía inversa (BLA-vHPC) puede provocar conductas similares a la ansiedad (Felix-Ortiz et al., 2013 ). Más bien, las manipulaciones optogenéticas de la vía vCA1-BA en nuestro estudio y otros trabajos recientes (Xu et al., 2016 ) demuestran que este camino es importante en las asociaciones de contexto-miedo. Por lo tanto, esta vía puede ser más especializada para codificar asociaciones de valencia de contexto aprendidas (en lugar de representaciones de valencia ansiogénicas innatas), por lo que la entrada de contexto de vCA1 a BA se emparejará con estímulos aversivos en el nivel de BA. Además, la actividad en las células de ansiedad vCA1-BA puede no ser suficiente para conducir el comportamiento de evitación a contextos innatos ansiogénicos en ausencia de una asociación aprendida de miedo a contexto.
Nuestros estudios plantean la posibilidad intrigante de que las subpoblaciones de neuronas vCA1 están programadas para responder a entornos que producen evitación innata, mientras que otras poblaciones pueden estar programadas para responder a entornos que provocan un enfoque. Esto podría lograrse mediante el enrutamiento selectivo de la información sensorial a las poblaciones de vCA1 que pueden dirigir directamente las respuestas de comportamiento positivas o negativas a través de sus flujos de dianas límbicas segregadas. Mientras que los proyectores vCA1-LHA están enriquecidos en células de valencia negativa, los proyectores vCA1-NAc pueden enriquecerse en células de valencia positiva ( Britt et al., 2012 , Ciocchi et al., 2015 , Okuyama et al., 2016 ). Este modelo sería similar al descrito en el BLA, donde distintas subpoblaciones de neuronas responden a la calidad de valencia positiva o negativa ( Belova et al., 2007 , Gore et al., 015, , Namburi et al., 2015 , Paton et al., 2006 , Uwano et al., 1995). Si vCA1 estuviera organizado de manera similar, predeciríamos que las neuronas vCA1 individuales responderían a diversos tipos de estímulos sensoriales de la misma valencia para impulsar respuestas conductuales análogas. Un enfoque para probar esta hipótesis será evaluar si las células de ansiedad vCA1 también responden a otros estímulos sensoriales aversivos innatamente, como olores aversivos o estímulos dolorosos. Es importante destacar que en el estudio actual utilizamos imágenes de calcio para probar las respuestas celulares a los comportamientos que se ejecutan en escalas de tiempo lentas (segundos), que son compatibles con las limitaciones temporales de la dinámica del calcio. Las futuras grabaciones electrofisiológicas de una sola unidad específicas de la vía tanto en la vHPC como en las regiones objetivo pueden dilucidar las respuestas a conductas más rápidas relacionadas con la ansiedad,
Nuestros hallazgos proporcionan información novedosa sobre la representación de información innatamente adversa en la vHPC y el papel de las proyecciones subcorticales de vCA1 específicas de la vía en la generación de conductas relacionadas con la ansiedad. La identificación de un nuevo circuito vCA1-LHA que controla rápidamente el comportamiento relacionado con la ansiedad, sin afectar el miedo aprendido, puede proporcionar nuevos objetivos para el tratamiento del trastorno del estado de ánimo y la ansiedad.
Cirugías Estereotácticas
Para todos los procedimientos quirúrgicos, los ratones se anestesiaron con isoflurano al 1,5% a una velocidad de flujo de oxígeno de 1 l / min y se fijaron en un marco estereotáctico (David Kopf, Tujunga, CA). Los ojos se lubricaron con una pomada oftálmica y la temperatura corporal se mantuvo a 37 ° C con un recirculador de agua tibia T / pump (Stryker, Kalamazoo, MI). Se afeitó el pelo y se esterilizó el sitio de la incisión antes de comenzar los procedimientos quirúrgicos, y se proporcionaron solución salina subcutánea y carpofeno de forma perioperatoria y durante 2 días después de la operación para prevenir la deshidratación y para la analgesia.
Para la obtención de imágenes de Ca 2+ in vivo , los ratones se sometieron a una única cirugía en la que se inyectaron unilateralmente 500 nl de virus GCaMP6f con una jeringa Nanoject (Drummond Scientific, Broomall, PA) antes de implantar una lente GRIN en el lugar de la inyección. Las lentes GRIN fueron implantadas con métodos descritos previamente (Resendez et al., 2016 ). Brevemente, se realizó una craneotomía centrada en el sitio de implantación de la lente, y la duramadre se extrajo de la superficie del cerebro y se limpió con una corriente de solución salina estéril y lanzas de absorción (Fine Science Tools (FST), Foster City, CA) antes de bajar el GRIN Lente (no se aspiró tejido fuera del sitio). Se insertaron 3 tornillos de cráneo (FST, Foster City, CA) en lugares espaciados uniformemente alrededor del sitio de implantación, y la lente se bajó lentamente en pasos DV de 0,1 mm y luego se fijó al cráneo con cemento dental (Dentsply Sinora, Filadelfia, PA) . Para la obtención de imágenes de vCA1, se usó una lente GRIN de ∼0.5 mm de diámetro, 6.1 mm de largo, y para dCA1 se usó una lente de GRIN de ∼1.0 mm de diámetro, 4 mm de largo (Inscopix, Palo Alto, CA). Las coordenadas de la inyección viral fueron (en mm, del tejido cerebral en el sitio): (vCA1: −3.16 AP, 3.25 ML, −3.85, −3.50, −3.25 DV; dCA1: −2.15 AP, 1.85 ML, −1.55, −1. 65 DV) y las coordenadas de la lente fueron (en mm, desde el cráneo hasta la craneotomía): (vCA1: −3.16 AP, 3.50 ML, −3.50 DV; dCA1: −2.15 AP, 1.30 ML, −1.30 DV). Al término de la cirugía, la lente se protegió con caucho de molde líquido (Smooth-On, Lower Macungie, PA) y los experimentos de imágenes comenzaron 3 semanas después.
Para las cirugías optogenéticas, los ratones se sometieron a una cirugía única en la que se inyectaron 500 nl de virus de opsina en la subregión vCA1 con una jeringa Nanoject como se describe anteriormente, antes de implantar fibra óptica en el sitio objetivo. Las fibras ópticas se realizaron con procedimientos previamente publicados ( Kheirbek et al., 2013 ), y se cortaron a ∼5mm de longitud para la implantación. Se implantó un solo tornillo de cráneo para permitir una mejor adherencia del cemento dental a la superficie del cráneo. El virus se inyectó en vCA1 en las siguientes coordenadas para todas las manipulaciones optogenéticas (en mm): (−3.16 AP, 3.30 ML, −3.85, −3.50, −3.00 DV desde el cerebro en la craneotomía). El silenciamiento del cuerpo celular vCA1 se realizó con un implante bilateral de fibra óptica y virus en las siguientes coordenadas (en mm): (−3.20 AP, 3.35 ML, −3.50 DV del cerebro en la craneotomía). La activación terminal de vCA1-BA se realizó bilateralmente con implante de fibra óptica a (en mm, desde el cerebro a la craneotomía): (−1.70 AP, 3.00 ML, −4.00 DV), y vCA1-LHA se realizó con un virus unilateral y fibra óptica implantada en : (−1.95 AP, 0.50 ML, −4.75 DV). Las cirugías de silenciamiento del cuerpo celular vCA1-LHA-ArchT se realizaron de forma bilateral con CAV2-Cre inyección en LHA en las coordenadas anteriores. Para el silenciamiento del cuerpo celular vCA1, se permitió que los ratones se recuperaran durante 4 semanas antes de comenzar los experimentos de comportamiento. Para la activación terminal, los experimentos comenzaron 8 semanas después de la cirugía para permitir una expresión viral suficiente y el tráfico de opsina a los terminales de axones.
Para los estudios retrógrados de CTB, se inyectaron 290nl de CTB conjugado (Life Technologies, Carlsbad, CA) unilateralmente en las subregiones LHA y BA o LHA y mPFC en una sola cirugía en las siguientes coordenadas (en mm del tejido cerebral en el sitio): (LH : −2.0 AP, 0.75 ML, −5.25, −5.0, −4.75 DV; BA: −1.70 AP, 3.0 ML, −4.25, −4.0 DV; mPFC: +1.90 AP, 0.3 ML, −2.75, −2.50 DV) y los ratones se perfundieron 7 días después de la inyección para histología.
Electrofisiología Patch-Clamp
Para las grabaciones de corte de terminal vCA1-ChR2, se anestesiaron ratones con expresión viral vCA1 de la opsina estimulante ChR2-eYFP (8 semanas después de la inyección viral, para permitir el tráfico de terminales de opsina a axón) mediante inhalación de halotano o isoflurano, descapitado y cerebro eliminado rápidamente. Se cortaron cortes coronales (350 μm) que contenían BA y LHA en un vibratome Leica VT1000S en solución de líquido cefalorraquídeo artificial de sacarosa parcial enfriada con hielo (ACSF) que contenía (en mM): 80 NaCl, 3.5 KCl, 4.5 MgSO 4 , 0.5 CaCl 2 , 1,25 H 2 PO 4, 25 NaHCO 3, 10 glucosa y 90 sacarosa equilibradas con 95% de O2 / 5% de CO2 y almacenadas en la misma solución a 37 ° C durante 30 minutos, luego a temperatura ambiente hasta su uso. Las grabaciones se realizaron a 30-32 ° C (TC324-B; Warner Instrument Corp) en ACSF (en mM: 124 NaCl, 2,5 KCl, 1 NaH 2 PO 4 , 25 NaHCO 3 , 20 glucosa, 1 MgCl 2 , 2 CaCl 2). Los terminales axones fluorescentes vCA1-ChR2-eYFP se ubicaron primero dentro de BA y LHA en un microscopio vertical Axioskop-2 FS (Zeiss). Las células rodeadas por estos axones se visualizaron luego a través de la óptica de contraste de interferencia de infrarrojo diferencial (IR-DIC) y se seleccionaron aleatoriamente para grabaciones de tensión de sujeción. Se utilizó una solución interna a base de cesio (en mM): 125 Cs-metanosulfonato, 4 NaCl, 10 HEPES, 1 EGTA, 4 MgATP, 0,3 Na 2.GTP, 10 Na-fosfocreatina, 5 QX 314-Cl). Las pipetas de parche se hicieron de vidrio de borosciliate (AM Systems) utilizando un extractor de micropipetas (Modelo P-1000; Sutter Instruments). En el baño, la resistencia inicial de la pipeta fue de 4.5-6.5 MΩ. Las grabaciones se realizaron sin corrección de potenciales de unión. Las señales de corriente y voltaje se registraron con un amplificador MultiClamp 700B (Molecular Devices, EE. UU.), Se digitalizaron a 5–10 kHz y se filtraron a 2,5–4 kHz. Los datos fueron adquiridos y analizados utilizando Axograph (Axograph Scientific, Sydney, Australia).
Para la estimulación óptica, se generaron pulsos de luz de 473 nm utilizando un láser DPSS de 100 mW (Opto Engine LLC, Midvale, UT) y se entregaron a través de un objetivo de 40X. Se utilizaron pulsos de luz individuales (1 ms de duración) cada 20 s para activar las fibras de vCA1 en el BA y el LHA mientras se registraban las EPSC monosinápticas evocadas por la luz en neuronas elegidas al azar.
Para los registros de cortes de cuerpos de células vCA1-ArchT, se anestesiaron ratones con expresión de ArchT (4 semanas después de la inyección viral) y se perfundieron con ACSF de sacarosa modificada que contiene (en mM) 75 NaCl, 2,5 KCl, 3,3 MgSO 4 , 0,5 CaCl 2 , 1NaH 2 PO 4 , 26.2 NaHCO 3, 22 glucosa, 52,6 sacarosa, 10 HEPES, 10 cloruro de colina, 1 piruvato, 1 L-ácido ascórbico (300 mOsml, pH 7,4). Se diseccionó el cerebro y se cortaron rodajas de 300 μm de grosor y se colocaron en una cámara de interfaz que contenía la misma solución modificada de sacarosa. Las rodajas se incubaron a 32 ° C durante 30 min, luego se mantuvieron a temperatura ambiente (23 ° C) en la cámara de interfaz durante al menos 1 h. Las grabaciones se realizaron a temperatura ambiente y se perfundieron con ACSF oxigenado que contenía (en mM) 119 NaCl, 2.5 KCl, 1.3 MgCl 2 , 2.5 CaCl 2 , 1.3 NaH 2PO 4 , 26.0 NaHCO 3 , 20 glucosa (∼300 mOsml) a 23 ° DO.
Los registros se hicieron a temperatura ambiente usando pipetas de parche tirados (5-7 mO) llenas con solución interna que contiene (en mM) 150 K-Gluconato, 1,5 MgCl 2 , 5,0 HEPES, 1 EGTA, 10 de fosfocreatina, 2,0 ATP, y 0,3 GTP. La luz verde se suministró a través de una lámpara de arco que pasó a través de un filtro de excitación TRITC y se entregó a través de un objetivo 40x centrado en el soma de la célula parcheada. Las grabaciones de patch-clamp se obtuvieron con los amplificadores de parche Multiclamp 700B, digitalizados con un Digidata 1322a, y los datos se recopilaron con el software pClamp 10 (Molecular Devices).
Ensayos de comportamiento
Elevated Plus Maze. Los ratones se colocaron en un laberinto de tamaño EPM estándar (13.5 «altura del laberinto del piso, 25» de longitud completa de cada tipo de brazo, ancho de brazo de 2 «, brazos cerrados de 7» de altura, con salientes de 0.5 «de alto / ancho en los brazos abiertos ), con lux650 lux ligero centrado sobre los brazos abiertos para promover la evitación. Los ratones se colocaron en la región central del laberinto y se les permitió explorar durante 10 minutos mientras se registraba el comportamiento con una cámara web EthoVision XT 10 (Noldus, Leesburg, VA) o una cámara digital (Carl Zeiss), y se analizaron con el software EthoVision o Software de seguimiento TopScan (Clever Sys, Reston, VA). Los comportamientos de Headdip en el EPM se puntuaron manualmente con el software Observer XT (Noldus, Leesburg, VA). Para los experimentos de silenciamiento de ArchT-GFP, los ratones se ejecutaron durante 20 minutos en el EPM para permitir un número suficiente de entradas de brazo abierto / eventos de activación con láser.
Nueva tarea de objetos . Los ratones se colocaron en una zona familiar (22 × 16 × 6 ”de largo, ancho y alto) que se les permitió explorar durante 20 minutos el día anterior, en condiciones de luz escasa (∼50 lux). El comportamiento durante la exposición inicial a la arena se registró y se siguió con el software EthoVision XT 10, y se dibujaron 4 zonas de esquina del mismo tamaño para determinar la preferencia de línea de base relativa para cada ubicación (6 × 5.5 ”de ancho y largo). Durante la sesión de objeto novedoso, se colocó un objeto novedoso que provocó un enfoque (un embudo tapado con cinta de color) en la zona de esquina menos preferida de la arena (desde el día 1 de seguimiento). A los ratones se les permitió explorar el terreno familiar durante 10 minutos y se registró el comportamiento con el software EthoVision XT 10 y la cámara web.
Prueba de campo abierto . Los ratones se colocaron en una arena (18 × 18 × 12 ”de largo-ancho-alto; Kinder Scientific, Poway, CA) con luz brillante (650 lux) centrada sobre la zona central, y se les permitió explorar durante 10 minutos mientras el comportamiento era Registrado y analizado con el software MotorMonitor.
Tarea de exploración de contexto para análisis de campo de lugar . Para el análisis de campo de lugar, a los ratones se les permitió explorar una arena novedosa (Contexto A- Contexto B- y Contexto A) (9,5 × 18 «de largo-ancho) durante 10 minutos cada uno en condiciones de luz baja lux, con un descanso de 10 minutos en una Transferencia de jaula entre sesiones. El contexto A era una arena lisa con paredes cortas (6 «de altura), mientras que el Contexto B se generó colocando camas estándar para el ratón, y paredes amarillas altas y redondeadas (10» altura) dentro de la misma arena que el Contexto A. La arena se mantuvo en el La misma ubicación para las 3 sesiones de imágenes, y el comportamiento se registró y siguió con el software EthoVision XT 10.
Preferencia de lugar en tiempo real . Los ratones se colocaron en una arena idéntica de 2 cámaras (18,5 × 10 × 8 ”de largo-ancho-alto) con camas de ratón estándar y lux de poca luz, y se les permitió explorar libremente ambas cámaras durante 20 minutos mientras se registraba el comportamiento con EthoVision XT 10 software.
Condicionamiento del miedo contextual. Los ratones se ejecutaron a través de un paradigma de condicionamiento del miedo contextual de 2 días. El día 1, los ratones se colocaron en una caja de choque de acondicionamiento de miedo estándar (Coulbourn Instruments, Holliston, MA) con las siguientes indicaciones contextuales: (olor a anís, ruido blanco y una luz encendida dentro de la cámara), y se les permitió explorar el Conéctelo durante 3 minutos antes de recibir un shock de pie de 2 s 0.7 mA con fuerza. En el día 2, los ratones se colocaron de nuevo en el mismo contexto durante 3 minutos para evaluar la congelación durante la recuperación de CFC. Para la modulación del terminal vCA1-LHA, al final de la recuperación del día 2, a los ratones se les aplicó un choque de pies adicional de 2 s para volver a entrenarlos para las pruebas de recuperación del día 3. Comportamiento fue la grabación con el software de video FreezeFrame (Coulbourn Instruments, Holliston, MA),
Imágenes de Ca 2+ que se mueven libremente
3 semanas después de la cirugía, los ratones se examinaron para determinar la expresión de GCaMP con un microscopio miniaturizado (Inscopix, Palo Alto, CA) y los procedimientos descritos anteriormente (Resendez et al., 2016). Los ratones se anestesiaron brevemente con isoflurano al 1,5% a 1 l / min de flujo de oxígeno y se fijaron en un marco estereotáctico. Se retiró el molde protector de goma de la lente, y se fijó una placa de base magnética a un microscopio y se bajó sobre la lente GRIN implantada para evaluar el FOV para las neuronas GCaMP +. Si las neuronas de GCaMP + eran visibles, la placa de base se cementó dentalmente en su lugar en el casco del ratón para permitir la creación de imágenes del mismo FOV durante varias semanas. Una vez que se colocó la placa base, se usó el mismo microscopio para cada sesión de imágenes con ese mouse, y el plano focal en el hardware del miniscopio no se alteró en todos los experimentos de imágenes para garantizar un FOV constante en todas las sesiones. Las sesiones de imagen de comportamiento de vigilia comenzaron el día después de la placa base y los ratones se anestesiaron brevemente (< 5mins) para adjuntar el miniscopio a la placa base cada día de sesión de imágenes. Se dejó que los ratones se recuperaran de la anestesia durante 30 minutos antes de comenzar la obtención de imágenes.
Los videos de Ca 2+ se grabaron con el software de adquisición nVista (Inscopix, Palo Alto, CA) y se activaron con un pulso TTL de EthoVision XT 10 y el sistema de caja Noldus IO para permitir la adquisición simultánea de Ca 2+ y videos de comportamiento. Los videos de Ca 2+ se adquirieron a 15 cuadros por segundo con una exposición de 66.56 ms. Se seleccionó una potencia de LED óptima para cada ratón en función de la expresión GCaMP en el FOV (valores de píxeles), y se usaron las mismas configuraciones de LED para cada ratón a lo largo de la serie de sesiones de imágenes.
Manipulaciones optogenéticas
Los ratones fueron manipulados y habituados a cables adaptadores de fibra óptica durante 3 días antes de comenzar los experimentos de comportamiento. Para los experimentos de silenciamiento de ArchT-GFP, se suministraron ∼10 mW de luz constante a través de un láser de 52m m (Opto Engine, Midvale, UT; se usó un láser de 594 nm en los experimentos de silenciamiento de arco vCA1-BA) a fibras ópticas implantadas en cerebro de ratón usando un cable de conexión de fibra óptica como se describió anteriormente (Kheirbek et al., 2013). Para los experimentos ChR2-eYFP, se administraron ∼5-8mW de 5ms 10hz o 20hz de pulsos de luz mediante un láser de 473nm 100mW (Opto Engine, Midvale, UT), y el protocolo de suministro de luz se controló a través de un estimulador Master-8 (AMPI, Jerusalén, Israel). Para los experimentos de silenciamiento ArchT-GFP en bucle cerrado, se utilizaron el software EthoVision XT 10 y el sistema de caja Noldus IO para registrar el seguimiento en vivo de los ratones mientras exploraban las tareas de EPM, OFT y RTPP. El láser se activó cuando los ratones fueron seguidos en vivo en EthoVision en una zona de estimulación previamente dibujada (brazos abiertos para EPM, centro para OFT y una cámara seleccionada al azar para RTPP). Para los experimentos de ChR2-eYFP, las manipulaciones optogenéticas OFT se realizaron en períodos de láser de 3 minutos (encendido / apagado de luz). RTPP se ejecutó como se describe anteriormente. En CFC, en días de luz,
Histología y microscopía confocal y epifluorescente.
Para toda la histología, los ratones se perfundieron transcardialmente con paraformaldehído al 4% (peso / volumen) en solución de tampón fosfato 1X (PBS) y luego se extrajeron los cerebros y se fijaron posteriormente en PFA al 4% durante 24 horas, después de lo cual se transfirieron a 30% de solución de sacarosa en PBS durante 2 días. Los cerebros saturados con sacarosa se congelaron instantáneamente y se cortaron en secciones coronales de 50 µg de espesor en un criostato (Leica CM 3050S). Las secciones se incubaron con 1: 1000 Hoechst en PBS 1x (Invitrogen, Carlsbad, CA) durante 10 minutos para marcar los núcleos celulares, y se montaron y se cubrieron con un reactivo antifade ProLong Gold (Invitrogen, Carlsbad, CA). La expresión viral endógena de fluoróforos se utilizó en todas las preparaciones histológicas (no se requirió inmunomarcaje para visualizar fluoróforos). Las diapositivas de histología se tomaron en imágenes en un microscopio confocal (Leica TCS SP8) utilizando un objetivo de 10x o 20x,
Se determinaron las colocaciones apropiadas de lentes de GRIN y de fibra óptica fijando los cerebros con las tapas de cabeza y los cráneos intactos durante 1 semana en PFA al 4% para mejorar la claridad de las lentes de GRIN y los tractos de fibra óptica. Los cerebros se colocaron luego en una solución de sacarosa al 30% como se describió anteriormente, y se recogieron los cortes en los pocillos de cultivo individuales para mantener el orden de AP preciso de las secciones para las reconstrucciones de colocación de lentes / fibra óptica. Luego, las secciones se montaron en orden AP, y la ubicación en el fondo de las puntas de fibra óptica y las lentes GRIN se determinaron visualmente para cada ratón mediante la inspección de las secciones en un microscopio epifluorescente.
Para los estudios retrógrados de CTB, las imágenes en mosaico se capturaron en un microscopio confocal con un objetivo de 10x, y las células rojas, verdes y amarillas se contaron con una caja de herramientas Contador de células ImageJ. La laminación de las neuronas marcadas con CTB se determinó midiendo la distancia de las células contadas al borde Pyr / Rad en ImageJ.
Para las mediciones de fluorescencia terminal anterógrada, los campos terminales vCA1 en BA y LHA se visualizaron en la misma ubicación AP (−1.70 mm) en ratones que expresan vCA1-ChR2-eYFP (8 semanas de inyección post-viral para permitir un tráfico suficiente de opsina para terminales de axon). Luego se tomaron imágenes de los subcampos BA y LHA en la misma sección con tiempos de exposición idénticos utilizando un objetivo de 2.5x en un microscopio epifluorescente vertical. Esto nos permitió controlar las diferencias en la línea base de fluorescencia de fondo entre las secciones, ya que las imágenes de los terminales BA y LHA se tomaron de las mismas secciones, y los niveles de fluorescencia se compararon en forma de pares. Las ROI de BA y LHA se dibujaron a mano en ImageJ, y se usó el valor de fluorescencia promedio para las comparaciones.
Procesamiento de imágenes
Para el procesamiento de imágenes de toda la población (promotor de Synapsin), el procesamiento de imágenes se realizó con el software Mosaic (versión 1.0.5b; Inscopix, Palo Alto, CA). Los videos se muestrearon en forma descendente con un factor de agrupación de 4 (16x), y el movimiento lateral del cerebro se corrigió mediante el motor de registro Turboreg (Ghosh et al., 2011, Ziv et al., 2013) que utiliza un cuadro de referencia único y funciones de alto contraste en la imagen para cambiar los cuadros con movimiento a las posiciones XY coincidentes en todo el video. Se recortaron los bordes negros de la conversión XY en la corrección de movimiento y se detectaron cambios en la fluorescencia al generar un video ΔF / F o usando una imagen de proyección z mínima de toda la película como la referencia F o para normalizar las señales de fluorescencia a la fluorescencia mínima de píxeles dentro del marco. Luego, los videos se muestrearon temporalmente con un factor de binning de 3 (hasta 5 cuadros por segundo). Las putativas células individuales y las señales de Ca 2+ se aislaron con un algoritmo automatizado de segmentación celular que emplea análisis de componentes principales e independientes en videos de ΔF / F o (Mukamel et al., 2009). Las supuestas células identificadas se clasificaron luego mediante inspección visual para seleccionar las unidades con la configuración espacial apropiada y la dinámica de Ca 2+ coherente con las señales de las neuronas individuales. Los eventos transitorios de Ca 2+ se definieron luego mediante un algoritmo de detección de eventos Ca 2+ que identifica picos de gran amplitud con tiempos de aumento rápidos y decaimientos exponenciales (parámetros: tau = 200 ms, tamaño mínimo del evento transitorio de Ca 2+ = desviación media de la media).
Para imágenes específicas de proyección, los videos se agruparon espacialmente, se corrigieron con movimiento y se recortaron usando el software Mosaic de la misma manera que se describió anteriormente. Sin embargo, la segmentación de células se realizó utilizando un algoritmo automatizado mejorado optimizado para la obtención de imágenes de Ca 2+microendoscópica. para datos microEndoscópicos (CNMF-E) ( Zhou et al., 2016). CNMF-E se basa en un marco de factorización matricial no negativa restringida (NMF) ( Pnevmatikakis et al., 2016 ), y es capaz de lograr la eliminación de ruido, la desconvolución y la mezcla simultánea de estos datos de imágenes. A nivel técnico, utiliza un modelo novedoso para estimar y restar eficientemente las grandes señales de fondo presentes en los datos (los detalles del método se pueden encontrar en ( Zhou et al., 2016 )). Los videos corregidos por movimiento y recortados se ejecutaron a través del algoritmo CNMF-E con un diámetro de celda estimado de 18 píxeles (medido en celdas visibles en imageJ). Las neuronas putativas se identificaron y se clasificaron por inspección visible para la configuración espacial apropiada y la dinámica de Ca 2+como se describió anteriormente, y las unidades putativas se fusionaron o dividieron manualmente si la configuración espacial no coincidía con las regiones de interés (ROI) dibujadas manualmente en el administrador de ImageJ ROI de la inspección visual de una versión videoF / F o del video. En el documento, informamos los rastros temporales no difuminados extraídos por CNMF-E como actividad temporal de las neuronas. Estas trazas se escalaron en z con un nivel de ruido gaussiano estimado, correspondiente a una versión escalada de ΔF / F o de cada neurona. Ca 2+los eventos transitorios se definieron con un algoritmo de detección de eventos similar al descrito anteriormente. Los transitorios se puntuaron en Z con la media calculada a partir de los puntos de tiempo que carecen de actividad de Ca 2+ (definidos como puntos de tiempo con valores de fluorescencia menores que el cuantil 0.50 de todos los valores de fluorescencia de todas las células en el FOV). Los eventos de Ca 2+ se definieron como transitorios que excedían una amplitud de 2 sd desde una línea de base de 0.5 sd, con una duración mínima (calculada por [-ln (A / Ao) / t_half] donde Ao = 0.5 y A = amplitud de ese transitorio; t_half para GCaMP6f fue de 200 ms, tomado de (Chen et al., 2013)) antes de volver a un nivel de base de 0.5 sd. Luego, se detectaron eventos adicionales de aumento transitorio de Ca 2+ dentro de los transitorios de Ca 2+ detectados que eran grandes y de pico múltiple utilizando la función findpeaks en MATLAB (Mathworks, Natick, MA) con los siguientes parámetros (MinPeakProminence = 1.5 sd, MinPeakDistance = 1 s ). Todos los transitorios de Ca 2+ detectados se inspeccionaron visiblemente en cada celda para verificar la precisión. El área bajo la curva se calculó para los transitorios identificados desde el inicio transitorio (en el momento en que el transitorio excedió el umbral de 0,5 sd) hasta el desplazamiento (en el momento en que la amplitud del transitorio regresó al umbral de base de 0,5 sd).
Para el seguimiento de celdas a través de múltiples sesiones de imágenes, los videos de varias sesiones se concatenaron en un solo video grande, y la corrección de movimiento se ejecutó en el video concatenado desde un solo cuadro de referencia para asegurar que la traducción XY del algoritmo de corrección de movimiento TurboReg ajustara todos los cuadros a el mismo lugar . La precisión del seguimiento celular y la estabilidad espacial a lo largo de los días de imágenes en el video concatenado segmentado se cuantificó. Brevemente (ampliando la descripción detallada en la leyenda de la figura para S3A-E), se seleccionó el umbral de correlación temporal transitoria de Ca 2+ para identificar pares emparejados temporalmente entre los grupos de video concatenado y segmentación de células de video individuales mediante inspección visual del Ca 2+ superpuesto transitorios entre pares de celdas con correlación máxima (a cada celda se le asignó una supuesta coincidencia con la celda en el segundo video con el que su valor de correlación transitoria era máximo). Esto dio lugar a una distribución bimodal de las correlaciones temporales de pares de células, con una división en Rho = 0,4 (todos los pares de células con transitorios que excedían de 0,4 de correlación se consideraron una coincidencia).
Cuantificación y análisis estadístico
Todos los parámetros estadísticos para análisis específicos se informan en las leyendas de las figuras del documento.
Ca 2+ análisis de datos
Los eventos transitorios de Ca 2+ y el comportamiento del ratón se analizaron con las funciones personalizadas de MATLAB (Mathworks, Natick, MA) para calcular la tasa de Ca 2+transitorios por celda mientras los ratones exploraron diferentes zonas y diferentes ubicaciones espaciales XY de la arena. La ocupación en diferentes zonas de arena se definió en el software EthoVision y se exportó como una salida lógica a 30 cuadros por segundo (la ubicación xy del punto central del mouse se rastreó en la arena definida por Ethovision para clasificar las muestras de comportamiento en zonas de arena específicas). Para el puntaje de comportamiento de headdip, se usó el software Observer XT (Noldus, Leesburg, VA) para puntuar manualmente los eventos de headdip y luego se exportó como una salida lógica similar al seguimiento de comportamiento de zona descrito anteriormente (se usó un sistema de puntaje de tecla de retención para registrar el cronometraje de eventos de headdip durante la duración de la sesión). Datos de comportamiento se muestrean a 5 cuadros por segundo para que coincida con Ca 2+ muestreo de datos transitorios, y Ca 2+los intervalos de tiempo transitorios se clasificaron en zonas de arena según el resultado del comportamiento de Ethovision emparejado en el tiempo para calcular el AUC y la tasa de transitorios de Ca 2+ en diferentes condiciones de comportamiento.
Para trazar ejemplos de mapas de calor de Ca 2+ por distancia de la actividad vCA1 en el EPM, se calculó el desplazamiento xy del punto central del ratón (desde el seguimiento de Ethovision) desde el punto central del EPM arena desde cada intervalo de tiempo de comportamiento, y los desplazamientos xy fueron categorizados por tipo de brazo y agrupados por distancia (contenedores de 0,5 cm). La media de la actividad transitoria de Ca 2+ (a partir de los transitorios identificados de Ca 2+ solamente) dentro de cada bandeja de desplazamiento xy de comportamiento se calculó y normalizó para cada celda individualmente. Para los mapas de calor de Ca 2+ por tiempo, la actividad transitoria media de Ca 2+ en todas las células en un FOV se calculó en intervalos de 200 ms.
Para definir la selectividad celular, los eventos de Ca 2+ se barajaron en el tiempo para celdas individuales (1000 iteraciones), y las tasas barajadas se volvieron a calcular en función de la categorización de la zona de tiempo de la zona de comportamiento como se describió anteriormente (las celdas de tiempo de comportamiento se mantuvieron constantes) para generar una distribución nula de las tasas de eventos de la zona Ca 2+ para cada celda. Una celda se consideró selectiva para una zona si su diferencia de tasa de eventos de Ca 2+entre zonas (EPM: abierto-cerrado; OFT: centro-periferia; Objeto nuevo: Zona de objeto nuevo-Zona neutral) excedió un umbral de 1SD desde la distribución nula ( el umbral se determinó comparando las tasas de zona para las "células neutrales" definidas en diferentes umbrales, y el umbral en el que las células neutrales no mostraron Ca 2+ significativo Se seleccionó tasa transitoria diferente entre zonas).
Colocar celula analisis
Los mapas de campo del lugar se dibujaron como se describió anteriormente (Leutgeb et al., 2007) con un tamaño de contenedor de ∼5cm 2 y un factor de suavizado sigma de 5, extraído solo de intervalos de tiempo móviles (los intervalos de tiempo inmóviles se definieron como velocidades <1cm / s, para episodios de comportamiento> 1 s de duración). Se requirió que se incluyeran en el análisis un mínimo de 10 eventos transitorios de Ca 2+ en la sesión, y los campos de lugar calificados se definieron como campos de lugar con 9 contenedores contiguos que contenían el 20% de la velocidad de disparo máxima. Los valores del contenido de información espacial se calcularon a partir de la primera sesión de imágenes de exposición del contexto-A. Los valores p de información espacial se calcularon como se describió anteriormente (Danielson et al., 2016a) generando una distribución nula de valores de contenido de información espacial por celda (barajando el tiempo de los eventos de Ca 2+ a tiempo para generar mapas de campo de lugar nulo, 1000 iteraciones), y comparando el valor verdadero del contenido de información espacial con esa distribución barajada. La estabilidad del campo del lugar se determinó comparando el valor R de las correlaciones del mapa del campo del lugar (correlación de Pearson). En este análisis solo se incluyeron las celdas con campos de lugar calificados en ambas condiciones (AA o AB).
Análisis de selectividad de tareas.
Para definir las neuronas selectivas a la tarea, las células se definieron por su selectividad para el brazo abierto EPM, la zona central OFT y la zona de objeto nuevo como se describió anteriormente. Luego se compararon poblaciones de células selectivas. La significación de la superposición se determinó mediante el muestreo aleatorio de una distribución simulada de células que contienen el porcentaje real de células de brazo abierto EPM en la población (muestra N = número total de células OFT de Novel Object en el centro, muestra simulada N = número total de todas las células incluidas en El análisis, muestreo a 10.000 iteraciones), para generar una distribución de superposición por muestreo aleatorio. El verdadero valor de superposición se calculó comparando el verdadero valor de superposición con estas distribuciones.
Referencias
• Adhikari A.
• Lerner TN
• Finkelstein J.
• Pak S.
• Jennings jh
• Davidson TJ
• Ferenczi E.
• Gunaydin LA
• Mirzabekov JJ
• Ye L.
• et al.
Doble disociación de la función dentro del hipocampo: memoria espacial e hiponeofagia.
Bannerman, DM, diácono, RMJ, Offen, S., Friswell, J., Grubb, M., Rawlins, JNP
Behavioral Neuroscience, Vol 116 (5), Oct 2002, 884-901
Las lesiones completas y dorsales del hipocampo perjudicaron el rendimiento espacial en 2 tareas de memoria de trabajo: alternancia recompensada en el laberinto en T y coincidencia con la posición en el laberinto de agua. En contraste, las lesiones del hipocampo ventral no tuvieron efecto en estas tareas, incluso cuando la dificultad de la tarea se incrementó por la introducción de retrasos. Las lesiones ventrales se parecían a las lesiones completas en la reducción de la ansiedad en 3 pruebas de ansiedad comúnmente usadas (interacción social, laberinto positivo e hiponeofagia). Las lesiones dorsales también parecieron ser ansiolíticas en la interacción social y en las pruebas del laberinto positivo, pero no afectaron la hiponeofagia. Las ratas con lesiones completas y dorsales mostraron hiperactividad, mientras que las ratas con lesiones ventrales no lo hicieron. Estos resultados muestran una doble disociación entre las lesiones del hipocampo dorsal y ventral (hiponeofagia vs. memoria espacial). Sugiriendo una diferenciación de la función a lo largo del eje septotemporal de esta estructura.
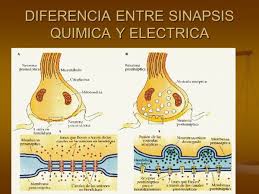
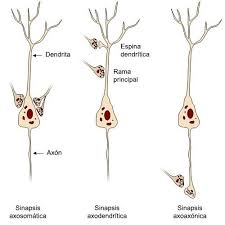 Sinapsis: qué son, tipos y funciones
Sinapsis: qué son, tipos y funciones


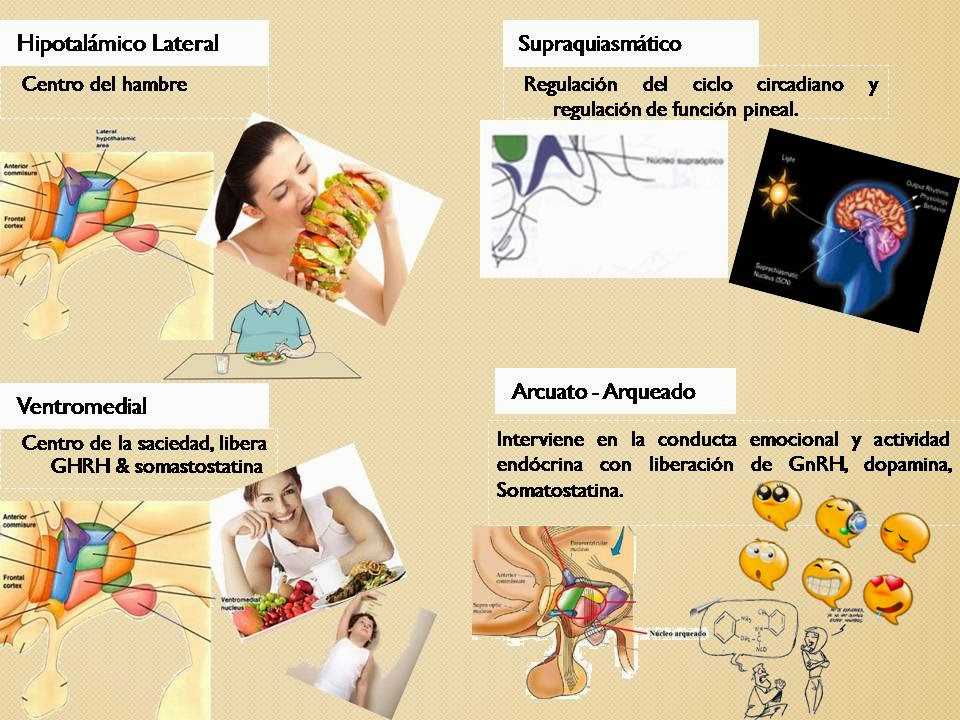 HIPOTALAMO Y FUNCIONES SUPERIORES
HIPOTALAMO Y FUNCIONES SUPERIORES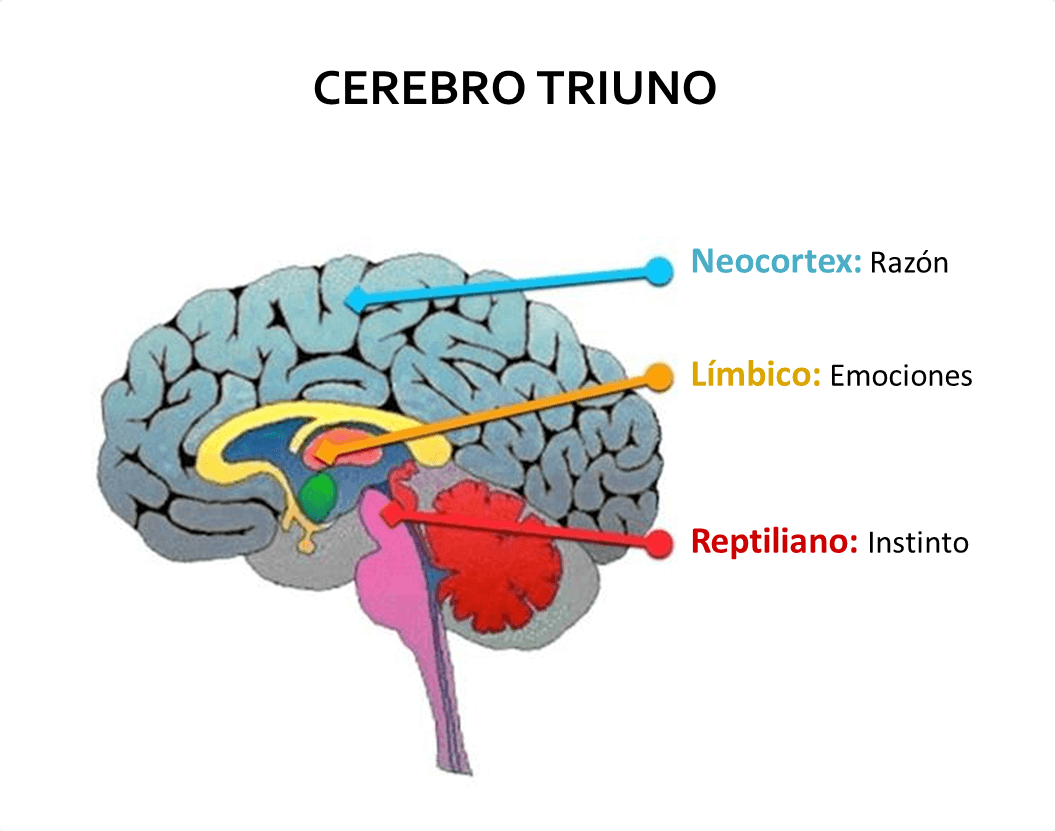 Las partes del encéfalo según Paul MacLean
Las partes del encéfalo según Paul MacLean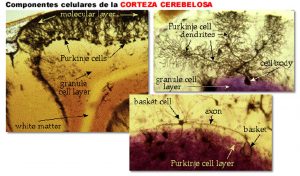
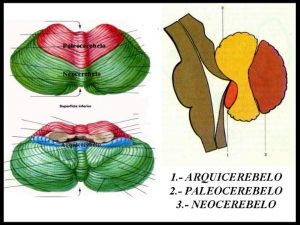 El cerebelo
El cerebelo