| ACTIVACIÓN E INHIBICIÓN DEL SISTEMA NERVIOSO | |

El concepto de inhibición es imprescindible para entender el funcionamiento de las funciones superiores concretamente de la memoria y de la tensión. Y también para la actividad motora y senitiva
Al igual que el arco reflejo , existen unos receptores, unos procesadores intermedos y unos efectores Todo esta soportado, por una anatomía, una bioquímica , y una respuesta , atención y memoria.
Al igual que el músculo para contraer el Bíceps el necesario una estructura intermedia que dé órdenes al Triceps, para que se relaje .
Toda esta complejidad fue ya estudiada por Charles Scott Sherrington, fue un médico neurofisiólogo británico, premio Nobel de Medicina, que estudió las funciones de la corteza cerebral.
Fue galardonado en 1932 con el premio Nobel de Medicina
y fue el primer autor que se preocupó de la inhibición del sistema nervioso. Este concepto estaba fuera del pensamiento de los científicos,. Era muy difícil introducir este concepto casi místico en la anatomía. Don Santiago, no se le ocurrió esta idea, porque era absolutamente somaticista en lo que al sistema nervioso se refiere.
Se tardo mucho tiempo en entender este mecanismo, de la acción y de la inhibición. Aunque era evidente, que para que actúe, un musculo efector, tiene que relajarse el musculo antagonista. Esta observación de Sherrington, se siguió del mecanismo a nivel molecular. La inhibición empieza a nivel celular, tras una química, muy compleja, donde los activadores, trabajan conjuntamente con los inhibidores y esto permite , no solo la motricidad, sino también la conducción nerviosa, y permite que el estimulo llegue a los múltiples receptores cerebrales, y provoque su efecto.
El término inhibición lateral es un fenómeno aplicable a una célula , cuando inhibe a otra adyacente para su, crecimiento o actividad. y diferenciación
Este fenómeno se lleva a cabo por la vía Notch, que media interacciones célula-célula. La célula inhibidora va a expresar: unas proteínas de membrana Delta, Jagged o Serrete. Estas van a ser ligando del receptor de membrana Notch, presente en las membranas de la mayoría de las células. Claude
Esta interacción provoca un cambio conformacional en Notch, que lleva al movimiento del dominio citoplasmático de esta, que es cortada por la proteasa Presennilina I. La porción escindida se transloca al núcleo, donde se va a unir a factores de transcripción de la familia CSL. Esta unión va a permitir que los CSL interaccionen con sus dianas génicas y las activen.
Se piensa que esta activación promueve el reclutamiento de histonacetiltransferasas, y con ello la descompactación de la cromatina. Las proteínas Notch son receptores muy importantes en el desarrollo del sistema nervioso, en el que esta vía es activada por la expresión de los genes proneurales, y llevan a la inhibición de determinadas células a neuronas
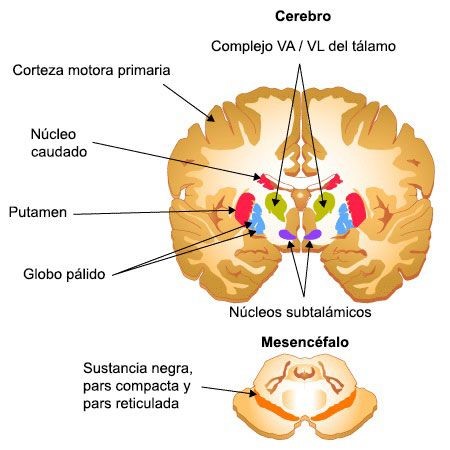
Anatomía y fisiología de los ganglios basales
Los ganglios basales son grandes estructuras neuronales subcorticales que forman un circuito de núcleos interconectados entre sí cuya función es la iniciación e integración del movimiento. Reciben información de la corteza cerebral y del tronco del encéfalo, la procesan y proyectan de nuevo a la corteza, al tronco y a la médula espinal para contribuir así a la coordinación del movimiento. Este circuito está compuesto por varias estructuras que se pueden categorizar según su anatomía o su función.
Anatómicamente los ganglios basales son masas de sustancia gris en el telencéfalo que incluyen: 1) núcleo caudado, 2) n lenticular (formado por el n putamen y el globo pálido externo e interno, 3) y la amígdala. Funcionalmente se relacionan a través de múltiples conexiones con núcleos próximos que incluyen al núcleo subtalámico (en el diencéfalo), la sustancia negra pars compacta y reticulata (en el mesencéfalo) y el n pedúnculopontino (en el puente).
El estriado es la estructura funcional «receptora» de aferencias extrínsecas a los ganglios basales, a través de diferentes neurotransmisores, en su mayoría excitatorios. Recibe proyecciones: 1) de la corteza cerebral (glutamatérgicas), 2) del tálamo (glutamatérgicas), y 3) de estructuras del tronco del encéfalo: SNpc (dopaminérgicas), del Núcleo pedúnculo pontino (NPP) del puente (glutamatérgicas y colinérgicas), del n dorsal del rafe (serotoninérgicas) y del locus coeruleus (noradrenérgicas).
La estructura eferente de los ganglios basales es el globo pálido interno, que envía proyecciones gabaérgicas para comunicarse con la corteza frontal a través los núcleos motores del tálamo (ventral anterior y ventrolateral). Las vías eferentes se dividen clásicamente en dos: la vía directa y la vía indirecta.
La vía directa se activa mediante los receptores dopaminérgicos tipo 1 (D1). Las neuronas espinosas medianas del estriado, producen una inhibición gabaérgica del GPI y la SNr que a su vez inhibe el tálamo cuya función es excitatoria sobre la corteza frontal. Por tanto, cuando el estriado recibe las proyecciones dopaminérgicas de la SNpc, se activa la vía directa y se activa la corteza motora (ya que se inhibe la proyección inhibitoria del GPI sobre el tálamo). La función de la vía indirecta es la contraria y normalmente está inhibida por las proyecciones dopaminérgicas de la SNpr a través de receptores dopaminérgicos D2. Al encenderse, a través de proyecciones gabaérgicas levanta el freno sobre el NST, cuya función habitual es la activación del GPI, que como se ha mencionado previamente, actúa como inhibidor tálamico y de la corteza.
En presencia de dopamina, neurotransmisor aferente fundamental de los ganglios basales, se activa la vía directa y por tanto la corteza está activada, mientras que se apaga la vía indirecta, y por tanto la corteza no esta inhibida.
Los trastornos de los ganglios basales se producen como consecuencia de la neurodegeneración o agresión secundaria de cualquiera de sus estructuras, produciendo un desequilibrio en este complicado circuito y por tanto una alteración de la coordinación motora. Se dividen en patologías hipocinéticas que implican pobreza de movimiento y en patologías hipercinéticas caracterizadas por exceso de movimiento
Los circuitos de formación de memoria se originan a partir de información procesada en áreas de asociación polimodal como la corteza frontal, temporal y parietal, de ahí el circuito lleva la información a la corteza parahipocámpica y corteza perirrinal y de ahí a la corteza entorrinal. Esta se comunica a través de la vía perforante con la circunvolución dentada, esta proyecta sus axones a través de las fibras musgosas a la región CA3 del hipocampo, que a su vez se conecta, con la vía colateral de Schaffer, a la región CA1 del hipocampo. Esta región se une con el subículo, el cual proyecta de vuelta a la corteza entorrinal. De aquí la información viaja hacia la corteza parahipocámpica y entorrinal y de ambas vuelve a las cortezas de asociación polimodal
El circuito consta de dos sub-circuitos: la vía directa y la vía indirecta. La sustancia negra compactada proyecta axones dopaminérgicos al putamen provocando la activación de este núcleo (cuando estimula los receptores D1), que aumenta su inhibición sobre el globo pálido interno y la sustancia gris reticulada mediante sus axones gabaérgicos. Esto produce una disminución de la actividad inhibitoria sobre el tálamo, el cual aumenta su activación sobre la corteza motora. De esta forma se comienza el movimiento. Este circuito es conocido como la vía directa Por otra parte, la sustancia negra compactada provoca la inhibición del putamen (cuando estimula los receptores D2), que disminuye su inhibición sobre el globo pálido externo, lo cual provoca un aumento de la actividad inhibitoria de este núcleo sobre el subtalámico, pero que al mismo tiempo este es excitado por las vías glutamatérgicas que vienen de la corteza, lo que permite que active al globo pálido externo y sustancia gris reticulada. Al ocurrir esto las vías inhibitorias de estos núcleos afectan al tálamo disminuyendo su activación, por lo cual ya no puede estimular la corteza motora y el movimiento se termina. Este circuito es conocido como la vía
Los procesos de memoria más estudiados son la habituación y la sensibilización. Un estímulo excitatorio que se repite produce una disminución del potencial sináptico de la neurona sensitiva sobre las interneuronas y sobre la neurona motora, lo que provoca que la respuesta disminuya. Esta disminución del potencial sináptico se produce por una disminución en la movilización de las vesículas que contienen el neurotransmisor glutamato, lo que provoca una menor liberación de la sustancia química y por ende disminuye la fuerza de la sinapsis, situación que puede durar varios minutos.
Este mecanismo es el que produce la memoria de corto plazo para la habituación. El sistema molecular de la memoria de corto plazo para la sensibilización es más complejo que el de la habituación.
Un estímulo nocivo aplicado en una vía produce un aumento de intensidad en otra vía a la cual se le aplica un estímulo no nocivo, esto mediante una interneurona facilitadora.
Existen dos vías de activación del botón terminal de la neurona sensitiva mediado por esta interneurona: a) en la primera vía la serotonina (5-HT) activa el receptor de la neurona sensitiva que a su vez activa una proteína G que aumenta la actividad de adenililciclasa que convierte el ATP en AMPciclico, el cual activa la proteincinasa dependiente de AMPc (PKA), esta fosforila los canales de potasio (k + ), esto prolonga el potencial de acción y permite más entrada de calcio (Ca ++ ) al botón terminal, lo que aumenta la liberación de glutamato (Glu) a la hendidura sináptica (Fig. 8.6); b) en la segunda vía la serotonina (5-HT) activa otro receptor de la neurona sensitiva que a su vez activa una proteína G que activa la fosfolipasa C (PLC) que a través del diacilglicerol, activa la proteincinasa C (PKC). Está en conjunto con PKA permiten la apertura de canales de Ca ++ con lo cual aumenta la liberación de Glu (Fig. 8.7). En ambos casos se produce una facilitación presináptica, ya que la interneurona facilitadora ayuda a la liberación del neurotransmisor de la neurona sensitiva ( …
Actualmente se acepta que la atención no es una función unitaria y que puede clasificarse en 2 grandes grupos: atención involuntaria y atención voluntaria. La primera corresponde al estado de alerta que ocurre desde que despertamos en la mañana y nos predispone para recibir los estímulos (Maureira & Flores, 2016); la atención voluntaria se clasifica en diversos tipos: a) atención focal, que ocurre cuando un sujeto presta atención a un solo estímulo, desechando otros (Maureira, 2018). También es llamada concentración (Ardila & Ostrosky, 2012); b) atención selectiva o capa- cidad de buscar un estímulo ignorando los demás; c) atención sostenida, definida como la capacidad de ejecutar una conducta cogni- tiva a lo largo de diversas ventanas tempora- les en contra de la fatiga (Maureira & Flores, 2016); d) atención alternante o capacidad de cambiar el foco atencional de un estímulo a otro rápidamente y; e) atención dividida, que corresponde a la capacidad de focalizar dos o más estímulos al mismo tiempo (Ardila & Ostrosky, 2012). …
La primera corresponde al estado de alerta que ocurre desde que despertamos en la mañana y nos predispone para recibir los estímulos (Maureira & Flores, 2016); la atención voluntaria se clasifica en diversos tipos: a) atención focal, que ocurre cuando un sujeto presta atención a un solo estímulo, desechando otros (Maureira, 2018). También es llamada concentración (Ardila & Ostrosky, 2012); b) atención selectiva o capa- cidad de buscar un estímulo ignorando los demás; c) atención sostenida, definida como la capacidad de ejecutar una conducta cogni- tiva a lo largo de diversas ventanas tempora- les en contra de la fatiga (Maureira & Flores, 2016); d) atención alternante o capacidad de cambiar el foco atencional de un estímulo a otro rápidamente y; e) atención dividida, que corresponde a la capacidad de focalizar dos o más estímulos al mismo tiempo (Ardila & Ostrosky, 2012). …
Se acepta que la atención no es un proceso único, sino que puede ser caracterizado como diversas funciones:
a) Estado de alerta, que corresponde a la atención involuntaria y sirve para aumentar la disposición para recibir información del entorno;
b) Atención selectiva, que corresponde a un tipo de atención voluntaria y sirve para seleccionar un estímulo específico ignorando los demás;
c) Atención sostenida, otro tipo de atención voluntaria, que corresponde a la capacidad de mantener una misma conducta a través del tiempo y la fatiga;
d) Atención alternante, un tipo de atención voluntaria, que se manifiesta como la capacidad de cambiar el foco de atención de un objeto a otro;
e) Atención dividida, otro tipo de atención voluntaria, que corresponde a la capacidad de focalizarse en dos o más estímulos al mismo tiempo (Maureira y Flores, 2016). El ejercicio físico puede mejorar los niveles de muchas funciones cognitivas, constituyéndose como una herramienta importante para potenciar la actividad cerebral
