CONCEPTO DE INTELIGENCIA ARTIFICIAL
La inteligencia artificial se suele definir como la capacidad que tienen artilugios artificiales de realizar tareas propias de una inteligencia humana
Fue acuñado en 1956 por John McCarthy lo definido como la ciencia y el ingenio de hacer máquinas inteligentes especialmente programas de cómputo

Las ideas más básicas sobre este concepto se remontan a los griegos de hecho Aristóteles fue el primero en definir un conjunto de reglas que describen una parte del funcionamiento de la mente para obtener conclusiones racionales por otra parte testigos de Alejandría fue el primero en construir una máquina auto controlada que fue un regulador de flujo de agua, varios siglos más tarde Alan Turing diseñó una máquina universal que demostraba la viabilidad de un dispositivo físico para implementar cualquier cómputo formalmente definido sin embargo a pesar de todos los avances que se han realizado desde entonces el concepto sigue estando en índices lo que está claro es que hay cuatro tipos de inteligencia artificial estos.
Son sistemas que actúan como un ser humano
sistemas que piensan cómo ser humanos
sistemas que piensan racionalmente y que imitan el procesamiento lógico delas personas
y sistemas que actúan racionalmente.
A su vez la inteligencia artificial se divide en dos escuelas la primera la convencional que consiste en un estudio del comportamiento humano la segunda la computacional que consiste en un aprendizaje interactivo.
La inteligencia artificial es la que se dedica a la investigación científica con ayuda de un ordenador y más aún con un conjunto de ellos podemos dar respuestas a todas esas cuestiones que nos llevamos preguntando durante todos estos años pero aún así esta inteligencia artificial no busca comprender cómo funciona nuestro cerebro sino que imita alguna de sus funciones
Una inteligencia artificial no tiene por qué parecerse del todo a una inteligencia humana.
La definición habitual significa eso , las tareas propias de una inteligencia humana
Si consideramos la capacidad de cálculo entonces eso si las máquinas la tienen
Si consideramos la capacidad de memorizar datos pues también
Conforme los ordenadores fueron siendo más capaces también se fueron atreviendo con cosas más de humanos inteligentes, juegos complicados como el ajedrez y otros para los que podían crear ciertas estrategias apoyadas sobre todo en su capacidad de cálculo y de memoria y poco a poco se iba avanzando en el terreno de la inteligencia que las matemáticas iban logrando
Pero hay cosas demasiado humanas que quedaban fuera del alcance de la inteligencia artificial como por ejemplo la capacidad de aprendizaje la creatividad o la autoconciencia, la inteligencia humana es probable más que esas tres cosas, pero si una máquina las logra por ahora no está nada mal.
La autoconciencia de momento está lejos, ahora mismo sólo la encontramos en las pelis en los videojuegos en las novelas o en los cómics.
Lo de la creatividad es más dudoso hay algoritmos a los que podemos empezar a atribuir ciertas formas de creatividad y es un terreno en el que se está trabajando mucho y hay ordenadores que son capaces de producir arte creativo o de inventarse chistes
Pero el 90% o más de las ocasiones en las que oyes hablar de que una inteligencia artificial ha hecho tal o cual cosa normalmente estamos en el terreno del aprendizaje y eso es un tema en el que hay avances espectaculares se llama aprendizaje automático o Machine learning
El Machin learning es es un conjunto de técnicas mediante las cuales un algoritmo tiene que realizar una tarea capaz de modificar su propio comportamiento basándose en los datos de que dispone o en lo bien o mal que lo haya hecho en el pasado o en lo que le digan otros que están bien o mal hecho
Osea aprender , esos algoritmos son capaces de aprender de sus errores , de algoritmos de aprendizaje automático que nos rodean cada día que son cada vez mejores y que son un tema de estudio.
Hay varios tipos:
Hay unos que se llaman de aprendizaje supervisado se les envía en un montón de datos que se llaman etiquetados o sea para los que se sabe la solución al problema que se les plantea y a medida que van procesando esos datos van aprendiendo.
A este proceso se le llama entrenamiento un ejemplo típico de estos más típicos, puede ser un algoritmo al que entrenamos para que sepa distinguir una foto mía de una foto de cualquier otra persona , en unos cuantos cientos de miles estoy yo y en otras no y le digo en cuáles sí estoy y en cuáles no estoy.
El algoritmo se entrena con esas fotos y luego cuando le llega una foto nueva, sabe si salgo en esa foto o no .
Estos algoritmos de aprendizaje supervisado se usan mucho y lo que necesitan son datos millones de datos etiquetados y tú les estás ayudando quizás sin saberlo sabes.
Para entrar en algunas webs que te dicen que Márquez fotos en las que salen semáforos o coches o peatones o señales o autobuses pues están etiquetando fotos que luego servirán para entrenar un algoritmo que reconozca esas cosas en imágenes y si se instaa en un coche autónomo o sea que haciendo bien lo de las fotos esas estás enseñando a conducir a los coches del futuro
Los problemas que resuelven estos algoritmos son super variados, un reconocimiento facial, reconocimiento de voz de huellas digitales coches automáticos.
En fin miles de cosas difíciles de imaginar.
Otro tipo de algoritmo de aprendizaje automático son los nuevos supervisados estos no entrenan como las anteriores con datos etiquetados estos se usan por ejemplo para agrupar datos que son parecidos entre sí i
Por ejemplo si el algoritmo agrupa a la gente que tienen los mismos gustos musicales o de ropa se podrían utilizar en publicidad y este no es su único uso hay algoritmos muy variados esto lo que hacen es definir una distancia entre datos por ejemplo entre tu historial de escuchas en spotify y el mío comparan nuestros gustos y así nos ofrecen canciones parecidas después se usan mucho también en aplicaciones científicas como en genómica por ejemplo.
Luego están los semi supervisados claro no va a haber supervisados no supervisados y ya está no aquí no somos binarios muy frente este lo que hace es que cuando tenemos pocos datos etiquetados por lo que sea consiguen unos pocos y usan el aprendizaje supervisado para etiquetar más datos por una parte y luego eso los usa en otro modelo de aprendizaje supervisado para resolver el problema que tengamos y finalmente otro tipo de aprendizaje automático muy usado es el aprendizaje por refuerzo este actúa por prueba y error y se usa mucho para aprender a jugar por ejemplo. En estos casos hay un concepto de recompensa que te dice cuando lo has hecho bien, por ejemplo ganar la partida , y así el algoritmo se queda con unos parámetros de una partida por ejemplo los movimientos que has hecho , las decisiones que ha tomado y si el resultado es que gana esos movimientos, los usará con más probabilidad en las próximas partidas y si le llevan a perder pues con menos probabilidad en aplicaciones científicas se usan muchísimo.
Para realizar estos tipos de aprendizaje hay muchas técnicas hay algoritmos específicos que se utilizan para cada problema en particular o que se mezclan para conseguir mejores modelos.
Entre los más famosos y utilizados estan los detalles están algunos que tienen que ver con estadística y probabilidad clásicas como algunos de reflexión regresión lineal regresión logística métodos etcétera Otros son árboles de decisión en los que se dirigen por una rama u otra según vas tomando decisiones Por ejemplo en Forest se generan varios árboles de decisión y compartes de los datos y se analiza el resultado de cada uno de ellos cuáles han sido las decisiones más comunes que vota la mayoría de los árboles etcétera es muy usado. No significa que mires neighbors y sirve por ejemplo para clasificar haciéndolo para cada en base a la clasificación de sus vecinos más próximos para ello hay que definir bien qué significa que los datos sean cercanos hay muchos otros support vector machine todos los degradan boosting .
Quizá la técnica más potente de todas son las redes neuronales que consisten en muchas pequeñas funciones matemáticas cada una de ellas llamada neurona o Zelda que se combinan entre sí se coordinan se pasan resultados unas a otras formando una red , cuando estas redes son grandes y con muchas capas s ellaman redes profundas y dan nombre a toda una rama del aprendizaje automático que se llama aprendizaje profundo o Deep learning que normalmente es aprendizaje supervisado
Usando redes profundas para entrenar al modelo.
Las aplicaciones son muchísimas algunas muy transformadoras hay algoritmos que hacen cosas increíbles Los de traducción automática que son cada vez mejores pero que también pueden cometer errores si no están bien diseñadas o incluso hacer cosas no tan buenas si no tenemos un poco de cuidado y sobre todo si no sabemos al menos algo de cómo funcionan están por todas partes
La inteligencia artificial y los algoritmos que hay detrás y sobre todo las matemáticas que les otorgan suponer
Bibliografia de
Internet









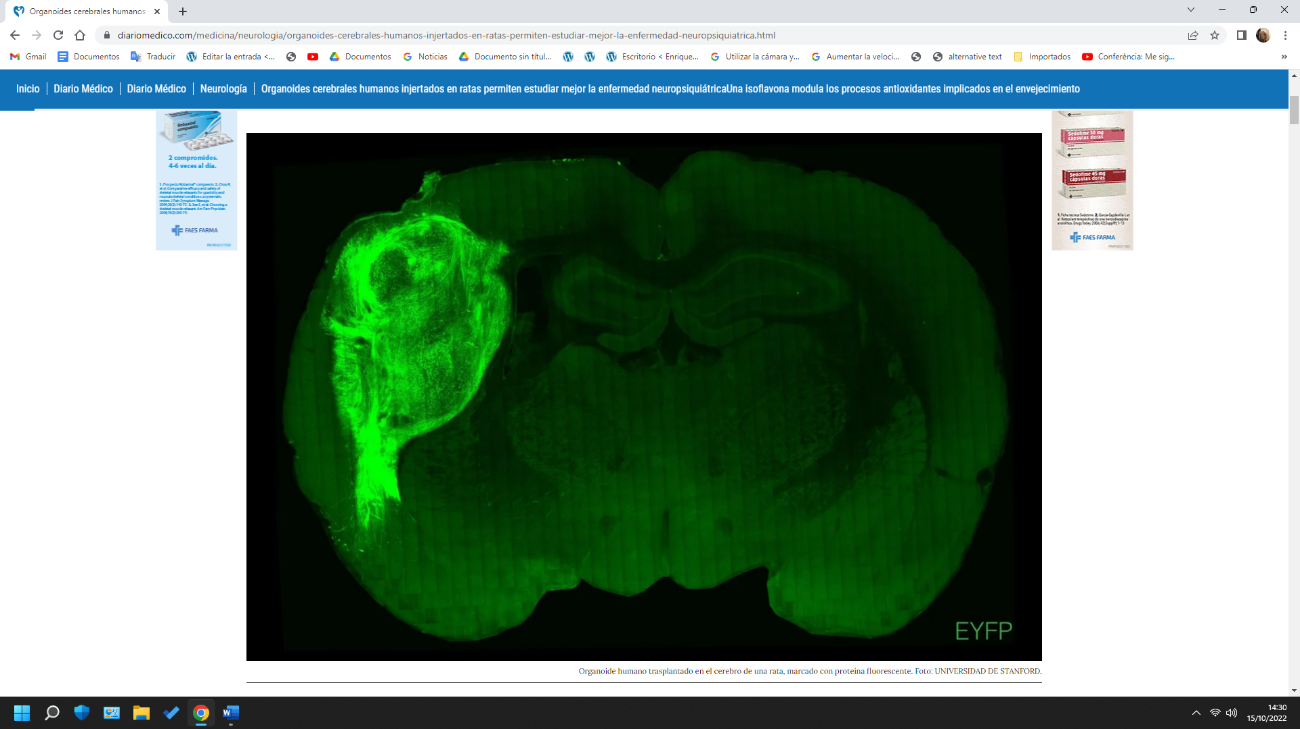
 Neuronas humanas derivadas de células madre y mantenidas ‘in vitro’ o trasplantadas en el córtex del cerebro de rata.
Neuronas humanas derivadas de células madre y mantenidas ‘in vitro’ o trasplantadas en el córtex del cerebro de rata.
 Los flavonoides de la soja pueden contrarrestar efectos antioxidantes en el organismo. Foto: DM.
Los flavonoides de la soja pueden contrarrestar efectos antioxidantes en el organismo. Foto: DM.
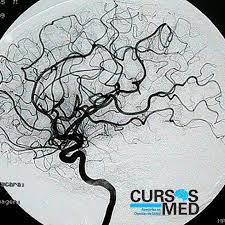
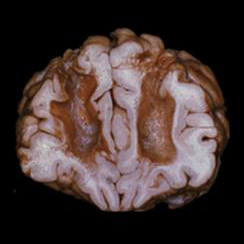
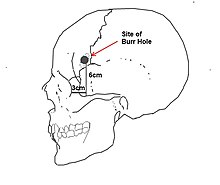 Punto de incisión normalizado para lobotomía pre-frontal / leucotomía desarrollada por Freeman y Watts.Este procedimiento se refiere comúnmente a toda clase de cirugías en los lóbulos frontales del cerebro, prefrontal —también llamada leucotomía—, frontal, temporal y parietal; sin embargo, debe llamarse propiamente lobotomía a la destrucción de las
Punto de incisión normalizado para lobotomía pre-frontal / leucotomía desarrollada por Freeman y Watts.Este procedimiento se refiere comúnmente a toda clase de cirugías en los lóbulos frontales del cerebro, prefrontal —también llamada leucotomía—, frontal, temporal y parietal; sin embargo, debe llamarse propiamente lobotomía a la destrucción de las